
4.Пути использования ацетил КоА. Механизм образования и значение ацетоуксусной кислоты. Биосинтез кетоновых тел. Кетоацидоз.
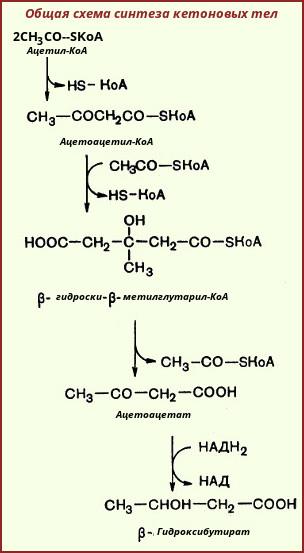
На первой стадии из двух молекул ацетил-КоА образуется ацетоацетил-КоА:

Реакция катализируется ферментом ацетил-КоА-ацетилтрансферазой (ацетоацетил-КоА-тиолазой). Затем ацетоацетил-КоА взаимодействует еще с одной молекулой ацетил-КоА. Реакция протекает под влиянием фермента гидроксиметилглутарил-КоА-синтазы:

Образовавшийся β-гидрокси-β-метилглутарил-КоА способен под действием гидроксиметилглутарил-КоА-лиазы расщепляться на ацетоацетат и ацетил-КоА:

Ацетоуксусная кислота способна восстанавливаться при участии НАД-зависимой D-β-гидроксибутиратдегидрогеназы; при этом образуется D-β-гидроксимасляная кислота (D-β-гидроксибутират). Следует еще раз подчеркнуть, что фермент специфичен по отношению к D-стереоизомеру и не действует на КоА-эфиры:

Существует также и второй путь синтеза кетоновых тел. Образовавшийся путем конденсации двух молекул ацетил-КоА ацетоацетил-КоА способен отщеплять коэнзим А и превращаться в свободную ацетоуксусную кислоту. Этот процесс катализируется ферментом ацетоацетил-КоА-гидролазой (деацилазой):

Однако второй путь образования ацетоуксусной кислоты не имеет существенного значения, так как активность деацилазы в печени низкая. В крови здорового человека кетоновые тела содержатся лишь в очень небольших концентрациях. При патологических состояниях (например, у лиц с тяжелой формой сахарного диабета, при голодании, а также у животных с острым аллоксановым диабетом) концентрация кетоновых тел в крови увеличивается и может достигать 20 ммоль. Такое состояние, которое получило название кетоза, возникает в тех случаях, когда скорость образования кетоновых тел превышает способность периферических тканей их утилизировать.
В последние годы начали постепенно накапливаться данные, указывающие на важную роль кетоновых тел в поддержании энергетического гомеостаза. Кетоновые тела — своего рода поставщики топлива для мышц, мозга, почек и действуют, возможно, как часть регуляторного механизма с обратной связью, предотвращая чрезвычайную мобилизацию жирных кислот из жировых депо. Печень в этом смысле является исключением, она не использует кетоновые тела в качестве энергетического материала.
В периферических тканях β-гидроксимасляная кислота окисляется до ацетоуксусной кислоты, а последняя активируется с образованием соответствующего КоА-эфира (ацетоацетил-КоА).
Существует два ферментативных механизма активации ацетоуксусной кислоты. Первый путь -это использование АТФ и HS-КоА аналогично тому, как при активации жирных кислот:

Реакция катализируется ферментом ацил-КоА-синтетазой, или тиокиназой. Второй путь активации — это перенос КоА от сукцинил-КоА на ацетоуксусную кислоту:

Данная реакция катализируется ферментом сукцинил-КоА-ацетоацетат-трансферазой. Возможно, что биологически более важным является именно этот путь активации ацетоацетата.
Образовавшийся в ходе этих реакций ацетоацетил-КоА подвергается далее тиолитическому расщеплению с образованием двух молекул ацетил-КоА:

А уже ацетил-КоА, как это известно, окисляется в цикле трикарбоновых кислот (цикл Кребса) до СО2 и Н20.
Кетоацидоз
В норме концентрация КТ в крови составляет 1—3 мг/дл (до 0,2 мМоль/л), но при голодании значительно увеличивается. Увеличение концентрации КТ в крови называют кетонемией. При кетонемии развивается кетонурия — выделение КТ с мочой. Накопление КТ в организме приводит к кетоацидозу, так как КТ (кроме ацетона) являются водорастворимыми органическими кислотами (рК
Ацидоз достигает опасных величин при сахарном диабете, так как концентрация КТ при этом заболевании может доходить до 400—500 мг/дл. Тяжёлая форма ацидоза — одна из основных причин смерти при сахарном диабете.
Биосинтез триацилглицеринов: стадии процесса. Липотропные факторы и их роль в биосинтезе липидов в печени.
Биосинтез триацилглицеринов. Образующиеся в результате биосинтеза жирные кислоты в организмах животных и человека в свободном виде встре- чаются лишь в незначительных количествах, а присутствуют главным образом в виде триацилглицеринов. Синтез триацилглицеринов происходит в печени и жировой ткани из КоА-производных жирных кислот через фосфатидную ки- слоту по реакции:
2 R C SКоА + О Н2С ОН НС Н2С О РО3Н2 ОН Н2С О СH Н2С О РО3Н2 С R O R C O O + 2 HS-KoA Фосфатидная кислота Фосфорилирование глицерина осуществляется глицеролкиназой за счет энергии АТФ. Глицерол-3-фосфат может образовываться и при восстановлении диоксиацетонфосфата. Гидролиз фосфатидной кислоты фосфатазой приводит к образованию 1,2-диацилглицерина, который, реагируя с другой молекулой ацил-КоА, обра- зует нейтральный триацилглицерин. В слизистой кишечника триацилглицерины синтезируются из свободных кислот, моно- и диацилглицеринов, но эти процессы характерны только для слизистой оболочки кишечника. Перенос остатка жирной кислоты происходит через ацильное производное КоА.
Липотропные факторы – вещества, способствующие синтезу фосфолипидов и препятствующие отложению триацилглицеролов в тканях.
Липотропный эффект этих соединений связан с тем, что общим предшественником триацилглицеролов и фосфолипидов является фосфатидная кислота. При недостатке липотропных факторов фосфатидная кислота используется преимущественно для синтеза триацилглицеролов. Они нерастворимы в воде и накапливаются в клетках, способствуя их жировому перерождению. Запомните, какие липотропные факторы можно использовать в качестве препаратов для его предупреждения:
Читайте также: Ткань для печати рисунка
1) холин – самое распространённое азотистое основание фосфолипидов;
2) метионин – незаменимая аминокислота, предшественник S-аденозилметионина, универсального донора метильных групп;
3) фолиевая кислота и витамин В12 – предшественники коферментов, участвующих в реакциях переноса СН3-групп.
При введении липотропных факторов снижается синтез триацилглицеролов и усиливается выведение жира из клеток при участии липопротеиновых комплексов
Acetyl-CoA
Acetyl CoA is a cofactor for pyruvate carboxylase, the first enzyme in the gluconeogenic pathway;
Related terms:
Lipids I
Source and Transport of Acetyl-CoA
Acetyl-CoA is synthesized in mitochondria by a number of reactions: oxidative decarboxylation of pyruvate; catabolism of some amino acids (e.g., phenylalanine, tyrosine, leucine, lysine, and tryptophan); and β-oxidation of fatty acids (see earlier). Since acetyl-CoA cannot be transported directly across the inner mitochondrial membrane to the cytosol, its carbon atoms are transferred by two transport mechanisms:
Transport dependent on carnitine: Carnitine participates in the transport of long-chain acyl-CoA into the mitochondria and plays a similar role in the transport of acetyl-CoA out of mitochondria. However, carnitine acetyl transferases have a minor role in acetyl-CoA transport.
Cytosolic generation of acetyl-CoA (citrate shuttle): This pathway is shown in Figure 16.8 . Citrate synthesized from oxaloacetate and acetyl-CoA is transported from mitochondria to the cytosol via the tricarboxylate anion carrier system and cleaved to yield acetyl-CoA and oxaloacetate.

Figure 16.8 . Cytoplasmic generation of acetyl-CoA via citrate transport and related reactions. PPP=pentose phosphate pathway; FAS=fatty acid synthase; —⊖→=negative allosteric modifier; —⊕→=positive allosteric modifier.
Thus, citrate not only modulates the rate of fatty acid synthesis but also provides carbon atoms for the synthesis. The oxaloacetate formed from pyruvate may eventually be converted (via malate) to glucose by the gluconeogenic pathway. The glucose oxidized via the pentose phosphate pathway augments fatty acid synthesis by providing NADPH. Pyruvate generated from oxaloacetate can enter mitochondria and be converted to oxaloacetate, which is required for the formation of citrate.
Carbohydrate Metabolism and Its Diseases
b Tricarboxylic Acid Cycle
AcCoA formed by the oxidative decarboxylation of pyruvate also has a number of metabolic routes available. AcCoA occupies a central position in synthetic and in oxidative pathways as shown in Figure 3-8 . The oxidative pathway leading to the breakdown of AcCoA to CO2 and H2O follows a cyclical pathway that is the tricarboxylic acid (TCA) cycle, citric acid cycle, or the Kreb’s cycle. The major steps involved are given in Figure 3-9 . In a single turn of the cycle, a mole of AcCoA enters, 2 moles of CO2 are evolved, and a mole of OAA is regenerated. The regenerated OAA may then condense with another mole of AcCoA, and the cycle continues. Citric acid is a symmetrical molecule that behaves asymmetrically as shown in Figure 3-9 . Also, the CO2 that is evolved is derived from that portion of the molecule contributed by OAA during each turn of the cycle. The expected distribution of carbon atoms from AcCoA in one turn of the cycle is also given in Figure 3-9 . During one turn of the cycle, a randomization of carbon atoms occurs at the succinate level such that CO2 derived from the carboxyl group of acetate will be evolved during the next turn of the cycle.

Figure 3-9 . Tricarboxylic acid cycle. The pathway for the entry of propionate into the metabolic scheme is also included. The asterisks give the distribution of carbon in a single turn of the cycle starting with acetyl-CoA. Note the randomization of carbon atoms at the succinate step.
In the process, 3 moles of NAD + and a mole of a flavin nucleotide (FAD) are reduced, and a mole of ATP is generated as noted in Figure 3-9 . In animal tissues, there is a cytoplasmic NADP + -linked isocitric dehydrogenase (ICD), which is not associated with the mitochondrial NAD + -linked ICD or other enzymes of the TCA cycle. The NADP + -ICD is another enzyme used as an aid to diagnose liver disease.
Methods in Methane Metabolism, Part A
Cornelia Welte , Uwe Deppenmeier , in Methods in Enzymology , 2011
2.4.4 Preparation of CO-dehydrogenase/acetyl-CoA synthase
CODH/ACS can be purified from M. thermoacetica ATCC 39073, as described by Ragsdale et al. (1983) , with modifications as specified ( Welte et al., 2010a ). All steps should be carried out anaerobically.
Читайте также: Ткани классификация ассортимент признаки идентификации в таможенных целях
M. thermoacetica cells are lyzed (suspended in 50 mM Tris–HCl, pH 7.5, 2 mM Na-dithionite) by French press treatment.
The lysate is applied to a DEAE cellulose column and eluted with a step gradient of NaCl from 0.1 to 0.5 M, CODH/ACS elutes at 0.3 M NaCl. To screen for the CODH/ACS-containing fractions, the following enzymatic test is used: 700 μL K-phosphate buffer (40 mM, pH 7.0, 1 μg mL − 1 resazurin, and 5 mM dithioerythritol), 5 mM methyl viologen, and 5% CO in the headspace. The reaction can be monitored at 600 nm at room temperature.
The enzyme containing fractions are applied to a Q-sepharose anion exchange column (elution at
0.4 M NaCl) and a phenyl–sepharose hydrophobic interaction column (elution at
The fractions from the phenyl–sepharose column that contain CODH/ACS are concentrated and the buffer is exchanged into 50 mM Tris–HCl (pH 7.6) by ultrafiltration.
Lysosomal Degradation of Heparin and Heparan Sulfate
C Acetyltransferase
Acetyl-CoA :α-glucosaminide N-acetyltransferase is the only lysosomal enzyme known to be involved in making a bond rather than hydrolysing one. The enzyme catalyzes the acetylation of the glucosamine amino group resulting from the action of heparan N-sulfatase ( Fig. 1 ). This is a biosynthetic reaction in what is a cascade of degradative process occurring in the lysosome to reduce heparin and HS to monosaccharides and inorganic sulfate. Adding to the enigma of this enzyme is its presence in the lysosomal membrane. The enzyme has not yet been purified to homogeneity, but partial purification has revealed much about its activity. On the cytoplasmic side, at neutral pH, the enzyme self-acetylates by transferring an acetyl group from acetyl-CoA. On the lysosomal side, at acidic pH, the enzyme transfers the acetyl group to a glucosamine residue ( 97 ). Evidence has also been provided that acetyl-CoA: α-glucosaminide N-acetyltransferase acts via a random-order ternary complex mechanism, which involves the utilization of cytosolic acetyl-CoA to transfer acetyl groups on to the terminal glucosamine residues of HS ( 98 ). The cDNA encoding this transferase remains to be isolated, although a deficiency in the enzyme has been associated with MPS IIIC, another of the more rare MPS subtypes. Here also the storage substrate is restricted to HS and its endo-degradation products.
Acetate
Martin Kohlmeier , in Nutrient Metabolism , 2003
Endogenous production
The metabolism of carbohydrates, amino acids, and fatty acids generates several hundred grams of acetate per day, mainly as acetyl-CoA . Depending on intakes, significant amounts of free actetate may also be generated from ethanol. Most is utilized within the cells or tissues where the acetate or acetyl-CoA is generated, some is transported to other tissues and utilized there.
Carbohydrates: The amount of acetate generated from glucose depends on the proportion used for glycolysis (as opposed to the smaller fraction metabolized via the pentose phosphate pathway) and the proportion used for the generation of oxaloacetate from pyruvate. Typically, about half a gram of acetate (as acetyl-CoA) is generated per gram of absorbed carbohydrate.
Amino acids: Acetyl-CoA is generated during the catabolism of isoleucine, leucine, and threonine. Lysine and tryptophan each generate two acetyl-CoA molecules. Metabolism of cysteine, alanine, and tryptophan generates pyruvate, which may be converted into acetyl-CoA. Acetoacetate is generated by the catabolism of phenylalanine, tyrosine, and leucine (for the latter in addition to one mole of acetyl-CoA). The acetoacetate can be activated by 3-oxoacid CoA-transferase (succinyl-CoA transferase, EC2.8.3.5) and then cleaved by acetyl-CoA C-acetyltransferase (thiolase, EC2.3.1.9) to generate two moles of acetyl-CoA. A minor pathway of threonine breakdown generates free acetate.

Figure 6.22 . Acetyl-CoA is a critical intermediate of fuel metabolism

Figure 6.23 . Endogenous sources ofacetate and its metabolites
Fatty acids: One mole of acetyl-CoA is released with each cycle of fatty acid beta-oxidation.
Alcohol: Ethanol is oxidized by various alcohol dehydrogenases (EC1.1.1.1) or the microsomal ethanol oxidizing system (MEOS, unspecific monooxygenases of the cytochrome P-450 family, EC1.14.14.1) in conjunction with several types of aldehyde dehydrogenases (EC1.2.1.3, EC1.2.1.4. and EC1.2.1.5) or acetaldehyde oxidase (EC1.2.3.1). Ethanol metabolism occurs mainly in the liver, and most of the resulting acetate is released into circulation ( Siler et al., 1999 ). One gram of ethanol generates about 1.3 g of acetate.
Читайте также: Блестящие полоски ткани 6 букв
Fiber: Normal intestinal bacteria break down non-digestible carbohydrates and release significant amounts of short-chain fatty acids including acetate.
Acetyl-Coenzyme A UV-Spectrophotometric Assay
Other Methods for Determination of Acetyl-CoA
Acetyl-CoA can be determined with CS alone according to equation (1); in this case the decrease of extinction of the thiol ester at 232 nm 1, 4, 9 or the formation of free SH groups (e.g. with nitroprusside 10 ) is measured. Neither method is suitable for measurements on tissue extracts or samples which contain much UV absorbing material or mercaptans. In the spectrophotometric method particular care should be taken with regard to the maintenance of an exact pH, because the extinction coefficients of acetyl-CoA and oxaloacetate are pH-dependent 1 .
The assay of acetyl-CoA by enzymatic acetylation of aromatic amines has a low substrate specifity (numerous other acyl thiolesters react). Arylamine acetylase is prepared from pigeon liver acetone powder 11 ; no commercial preparations are available. In addition acetyl-CoA can be determined by arsenolysis with phosphotransacetylase from Clostridium kluyveri; the assay is based on the measurement of the decrease in extinction at 232 nm 9 . This method is the most specific. Its main disadvantage lies in the difficulty of carrying out measurements on samples which have a strong UV absorption.
A modification of this method has been described by Garland et al. 12 , in which the CoA liberated by the arsenolysis is determined specifically by the use of 2-oxoglutarate dehydrogenase (2-oxoglutarate: lipoate-oxidoreductase (acceptor acylating), EC 1.2.4.2). This method combines the specificity of phosphotransacetylase with the measurement of NADH formation at 340 nm. 2-Oxoglutarate dehydrogenase is prepared according to Sanadi et al. 13 , and must be highly purified. Preparations of this enzyme are not available commercially.
The sensitivity of all the assay methods linked to the pyridine nucleotides can be increased by fluorimetry (see p. 1993). A radiochemical method for the determination of acetyl-CoA with CS in the presence of [ 14 C]-oxaloacetate has been described by Prinz et al. 14 (see p. 1994).
Under suitable conditions non-enzymatic methods can also be used. Examples of these methods are determination as the acetohydroxamate, the delayed nitroprusside reaction and UV spectroscopy 4 .
Pheromone Biosynthesis: Enzymatic Studies in Lepidoptera
DAVID MORSE , EDWARD MEIGHEN , in Pheromone Biochemistry , 1987
1 Reaction Pathways
Acetyl-CoA and malonyl-CoA are the basic building blocks used for the de novo biosynthesis of fatty acids in living organisms ( Volpe and Vagelos, 1973 , 1976 ). Acetyl-CoA normally arises in the cell as a product of the glycolytic cycle or by β-oxidation of fatty acids in the mitochondria. In addition, acetyl-CoA can also be formed by direct esterification of acetate in the cytoplasm by acetyl-CoA synthetases [reaction (1)]. The synthesis of malonyl-CoA is catalyzed by acetyl-CoA carboxylases [reaction (2)] which condense carbon dioxide with acetyl-CoA
in an ATP-dependent reaction. Fatty acids are then synthesized by condensation of two-carbon units from malonyl-CoA with a growing fatty acyl chain covalently linked to fatty acid synthase (X) in eukaryotic systems [reaction (3)] or
attached to the acyl carrier protein in prokaryotic systems; the release of carbon dioxide provides the driving force for the condensation reaction. The β-ketoacyl derivative is reduced, dehydrated, and reduced again to form the fatty acyl derivative two carbons longer [reaction (4)]. The cycle is then repeated with
condensation of another malonyl-CoA molecule with the elongated fatty acyl chain. Biosynthesis of palmitic acid by fatty acid synthase thus involves sequential condensation of seven malonyl-CoA residues with an acetyl-CoA primer and the oxidation of 14 molecules of NADPH [reaction (5)]. Seven different activities are involved in the reaction catalyzed by fatty acid synthases. These
activities are located on different polypeptides in a multienzyme complex in prokaryotic systems or in different domains on a single polypeptide chain of a multifunctional protein in eukaryotic systems ( Wakil et al., 1983 ).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом