
Глюкоза является основным метаболитом и транспортной формой углеводов в организме человека и животных. Источниками глюкозы являются углеводы пищи, гликоген тканей и процесс глюконеогенеза в печени и корковом веществе почек. Для включения глюкозы в метаболизм она должна фосфорилироваться с образованием глюкозо-6-фосфата (Г-6-Ф), который далее может превращаться по различным метаболическим путям. На Рис. 17.1. представлены основные пути метаболизма глюкозы.
Гликолиз – главный путь катаболизма глюкозы путем последовательных ферментативных превращений до лактата (без потребления кислорода – анаэробный гликолиз) или через окислительное декарбоксилирование пирувата до СО 2 и Н 2О (в присутствии кислорода – аэробный гликолиз).
Процесс аэробного гликолиза включает несколько стадий:
1. Аэробный гликолиз – процесс окисления глюкозы с образованием двух молекул пирувата;
2. Общий путь катаболизма, включающий окислительное декарбоксилирование пирувата до ацетил КоА и его дальнейшее окисление в цикле трикарбоновых кислот;
3. Цепь тканевого дыхания, сопряженная с реакциями дегидрирования, происходящими в процессе распада глюкозы.
Суммарный выход АТФ при окислении 1 моль глюкозы до СО 2 и Н 2О составляет 38 моль.
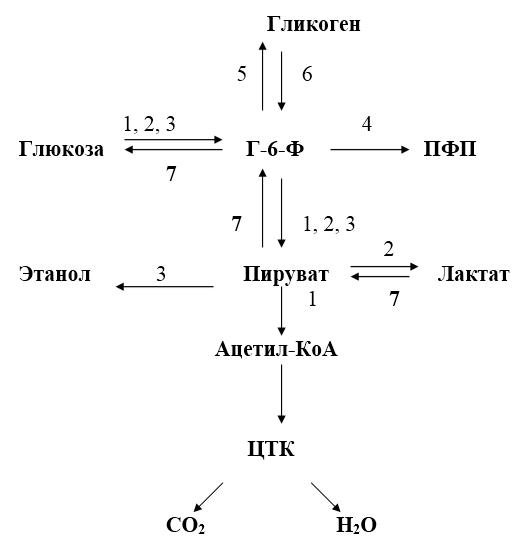
Рис. 17.-1. Общая схема путей метаболизма глюкозы.
1 – аэробный гликолиз; 2 – анаэробный гликолиз; 3 – спиртовое брожение; 4 – пентозофосфатный путь; 5 – синтез гликогена; 6 – распад гликогена; 7 – глюконеогенез.
Анаэробный гликолиз – процесс расщепления глюкозы с образованием в качестве конечного продукта лактата. Этот процесс протекает без использования кислорода и поэтому не зависит от работы митохондриательной сети. АТФ здесь образуется за счет реакций субстратного фосфорилирования. Баланс АТФ при анаэробном гликолизе составляет 2 моль в расчете на 1 моль глюкозы.
Аэробный гликолиз происходит во многих органах и тканях и служит основным, хотя и не единственным, источником энергии для жизнедеятельности.
Кроме энергетической функции гликолиз может выполнять и анаболические функции. Метаболиты гликолиза используются для синтеза новых соединений. Так, фруктозо-6-фосфат и глицеральдегид-3-фосфат участвуют в образовании рибозо-5-фосфата – структурного компонента нуклеотидов. 3-фосфоглицерат может включаться в синтез аминокислот, таких как серин, глицин, цистеин. В печени и жировой ткани ацетил-КоА, образующийся из пирувата, используется как субстрат при биосинтезе жирных кислот, холестерина.
Анаэробный гликолиз активизируется в мышцах при интенсивной мышечной работе, происходит в эритроцитах (в них отсутствуют митохондрии), а также в разных условиях ограниченного снабжения их кислородом (спазм и тромбоз сосудов, формирование атеросклеротических бляшек).
Данный текст является ознакомительным фрагментом.
Читайте также
Глава 4. Два пути
Глава 4. Два пути Факт или артефакт? Профессор С. Северин, узнав, что вслед за Шарфшвертом я освоил заокеанскую методику, попросил применить ее к другому объекту: вместо печени крысы надо было взять грудную мышцу голубя. План моего руководителя состоял в том, чтобы
ПУТИ СНАБЖЕНИЯ
ПУТИ СНАБЖЕНИЯ У берегов восточного Крыма, там, где горбится величественный горный массив Карадаг, прямо из голубых вод Черного моря поднимается грандиозная скала Золотые ворота, похожая на огромную арку, увенчанную шпилем. Старожилы окрестных городов и поселков,
7. Виды метаболизма бактерий
7. Виды метаболизма бактерий В процессе метаболизма выделяют два вида обмена:1) пластический (конструктивный):а) анаболизм (с затратами энергии);б) катаболизм (с выделением энергии);2) энергетический обмен (протекает в дыхательных
Читайте также: Пятновыводитель от шариковой ручки с ткани
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции Пер. А. НестеровойВ этой главе мы продолжим обсуждение эволюционной биологии в период до появления геномики. Многие из обсуждаемых направлений развития не являлись
Глава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы
Глава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы Пер. А. НеизвестногоВ предыдущей главе мы обсудили возможные сценарии возникновения клеток и (будем надеяться) достигли
Глава 4 Препятствия на пути кислорода
Глава 4 Препятствия на пути кислорода В нормальной атмосфере гемоглобин связывает только кислород. Это значит, что на связывание кислорода не оказывают воздействия другие компоненты воздуха: азот, двуокись углерода, пары воды или аргон. Гемоглобин собирает
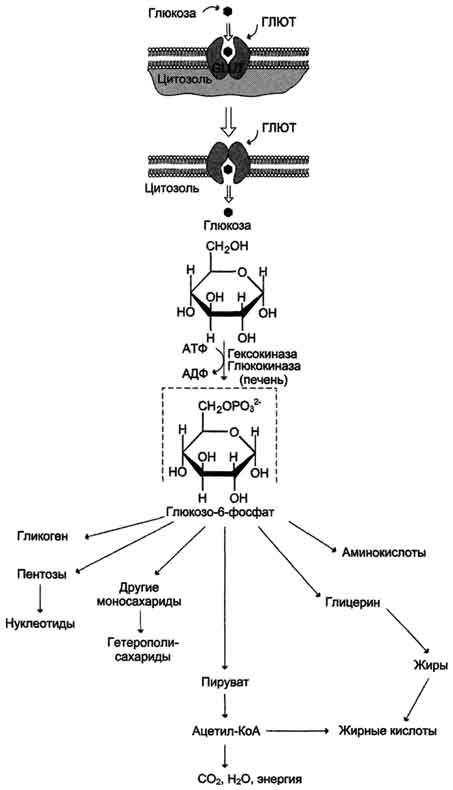
Транспорт глюкозы из крови в клетки
Транспорт глюкозы из крови в клетки Поглощение глюкозы клетками из кровотока происходит, также путем облегченной диффузии. Следовательно, скорость трансмембранного потока глюкозы зависит только от градиента ее концентрации. Исключение составляют клетки мышц и жировой
Глава 27. Регуляция и взаимосвязь метаболизма
Глава 27. Регуляция и взаимосвязь метаболизма Для нормального функционирования организма должна осуществляться точная регуляция потока метаболитов по анаболическим и катаболическим путям. Все сопутствующие химические процессы должны протекать со скоростями,
Взаимосвязь метаболизма
Взаимосвязь метаболизма Метаболизм в целом не следует понимать как сумму обменов белков, нуклеиновых кислот, углеводов и липидов. В результате взаимодействия обменов отдельных классов органических соединений возникает единая система метаболических процессов,
Особенности метаболизма в форменных элементах крови
Особенности метаболизма в форменных элементах крови Эритроциты:1. Зрелые эритроциты лишены ядра, поэтому в клетке не синтезируются белки. Эритроцит почти целиком заполнен гемоглобином.2. Эритроциты не имеют митохондрий, поэтому в клетке не протекают реакции ЦТК, ЦТД,
Глава 32. Особенности метаболизма в нервной ткани
Глава 32. Особенности метаболизма в нервной ткани Человеческий мозг – это самая сложная из всех известных живых структур. Нервной системе и, в первую очередь, головному мозгу принадлежит важнейшая роль в координации поведенческих, биохимических, физиологических
Концентрация глюкозы в крови – важнейшая константа
Концентрация глюкозы в крови – важнейшая константа На рис. 2.10 обмен углеводов представлен в виде схемы. Основным показателем состояния углеводного обмена является содержание глюкозы в крови. Нормальным является показатель от 4,4 до 6,6 миллимоля, что соответствует
Глава 24. ГДЕ СХОДЯТСЯ ВСЕ ПУТИ
Глава 24. ГДЕ СХОДЯТСЯ ВСЕ ПУТИ Последние четыре главы были посвящены, тем или иным образом, процессам, связанным с катаболизмом глюкозы — сначала до молочной кислоты путем анаэробного гликолиза, потом — до углекислоты и воды путем цикла Кребса. Однако нельзя сказать,
Глава IV. Пути изучения фенофонда
Глава IV. Пути изучения фенофонда Понятие «фенофонд» родилось в нашей стране. В московской школе генетиков, возглавляемой Н. К. Кольцовым, и в ленинградской школе, возглавляемой Ю. А. Филипченко, в 20-х годах интенсивно шло обсуждение популяционно-генетических вопросов. В
Глава 3. Неисповедимы пути полового отбора
Глава 3. Неисповедимы пути полового отбора Тайны полового отбора Эволюция в направлении производства половых клеток разного размера сама создает новое селективное давление, которое способствует дальнейшей дифференциации двух морфотипов гамет. Происходит это
Читайте также: Кофейные елки из ткани
Глюкоза как важнейший метаболит углеводного обмена.
Основным углеводом, участвующим в метаболизме, является глюкоза. Центральная роль молекулы глюкозы в углеводном обмене обусловлена:
1. Высокой растворимостью глюкозы.
2. Наличием реакционно-способной карбонильной группы в молекуле.
3. Оптимальной стабильностью пиранозного кольца.
Источники глюкозы для организма:
– животная и растительная пища;
– собственные резервные полисахариды (гликоген);
– другие моносахара (взаимопревращения);
– гликонеогенез (синтез глюкозы из неуглеводов). Пути расходования глюкозы:
– анаэробное и аэробное окисление в тканях – 60-65%;
– превращения в другие моносахара – 5-7%.
– дихотомический распад – анаэробный гликолиз и аэробное окисление;
– апотомическое окисление – пентозофосфатный путь.
Поскольку в составе основных углеводов пищи преобладает глюкоза, её можно считать основным продуктом переваривания углеводов. Другие моносахариды, поступающие из кишечника в процессе метаболизма, могут превращаться в глюкозу или продукты её метаболизма. Часть глюкозы в печени депонируется в виде гликогена, а другая часть через общий кровоток доставляется и используется разными тканями и органами.
Глюкоза является альдогексозой. Она может существовать в линейной и циклической формах. Циклическая форма глюкозы, предпочтительная в термодинамическом отношении, обусловливает химические свойства глюкозы. Как и все гексозы, глюкоза имеет 4 асимметричных углеродных атома, обусловливающих наличие сте-реоизомеров. Возможно образование 16 стереоизомеров, наиболее важные из которых D- и L-глюкоза. Эти типы изомеров зеркально отображают друг друга. Расположение Н- и ОН-групп относительно пятого углеродного атома определяет принадлежность глюкозы к D- или L-ряду. В организме млекопитающих моносахариды находятся в D-конфигурации, так как к этой форме глюкозы специфичны ферменты, катализирующие её превращения. В растворе при образовании циклической формы моносахарида образуются ещё 2 изомера (α- и β-изомеры), называемые аномерами, обозначающие определённую конформацию Н- и ОН-групп относительно С, У α-D-глюкозы ОН-группа располагается ниже плоскости кольца, а у β-D-глюкозы, наоборот, над плоскостью кольца.
Общая схема источников и путей расходования глюкозы в организме
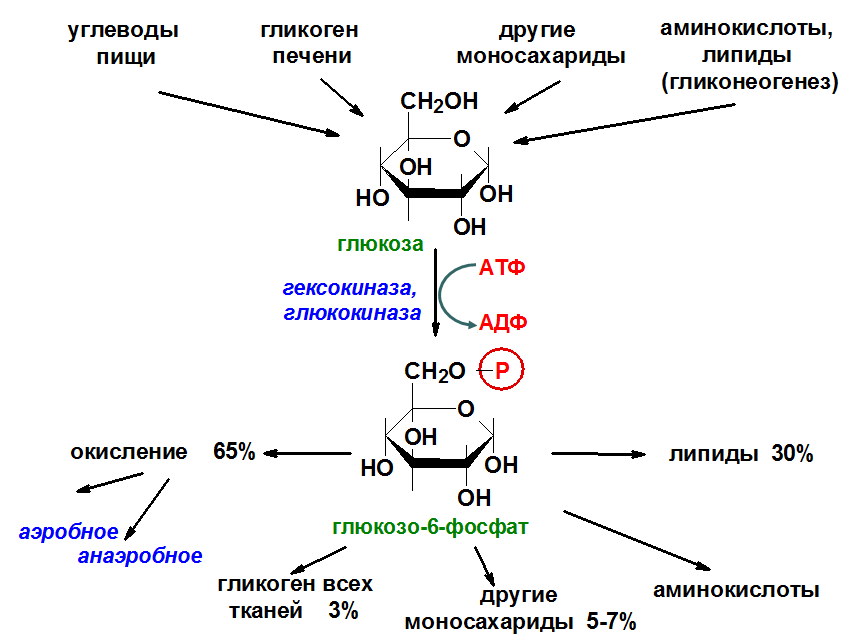
Ф ософорилирование глюкозы В дальнейших превращениях в клетках глюкоза и другие моносахариды участвуют только в виде фосфорных эфиров. Фосфорилирование свободных моносахаридов — обязательная реакция на пути их использования, она приводит к образованию более реакционно-способных соединений и поэтому может рассматриваться как реакция активации. Глюкоза, поступающая в клетки органов и тканей, сразу же подвергается фосфорилированию с использованием АТФ. Эту реакцию во многих тканях катализирует фермент гексокиназа, а в печени и поджелудочной железе — фермент глюкокиназа. Фосфорилирование глюкозы — практически необратимая реакция, так как она протекает с использованием значительного количества энергии. Образование глюкозо-6-фосфата в клетке — своеобразная «ловушка» для глюкозы, так как мембрана клетки непроницаема для фосфорилированной глюкозы (нет соответствующих транспортных белков). Кроме того, Фосфорилирование уменьшает концентрацию свободной глюкозы в цитоплазме(благоприятные условия для облегчённой диффузии глюкозы в клетки из крови)
Г люкокиназа. Фосфорилирование глюкозы в гепатоцитах в период пищеварения обеспечивается свойствами глюкокиназы. В этот период концентрация глюкозы в воротной вене больше, чем в других отделах кровяного русла и может превышать 10 ммоль/л, а следовательно, активность глюкокиназы в гепатоцитах повышается. Следует отметить, что активность глюкокиназы, в отличие от гексокиназы, не ингибируется продуктом катализируемой реакции — глюкозо-6-фосфатом. Это обстоятельство обеспечивает повышение концентрации глюкозы в клетке в фосфорилированной форме, соответственно её уровню в крови. Как уже упоминалось, глюкоза проникает в гепатоциты путём облегчённой диффузии при участии транспортёра ГЛЮТ-2 (независимого от инсулина). ГЛЮТ-2, так же, как глюкокиназа, имеет высокую Кm, что способствует повышению скорости поступления глюкозы в гепатоциты в период пищеварения, следовательно, ускоряет её фосфорилирование и дальнейшее использование для депонирования. Хотя инсулин и не влияет на транспорт глюкозы, он усиливает приток глюкозы в гепатоциты в период пищеварения косвенным путём, индуцируя синтез глюкокиназы и ускоряя тем самым Фосфорилирование глюкозы. Преимущественное потребление глюкозы гепатоцитами, обусловленное свойствами глюкокиназы, предотвращает чрезмерное повышение её концентрации в крови в абсорбтивном периоде. Это, в свою очередь, снижает последствия протекания нежелательных реакций с участием глюкозы, например гликозилирования белков.
Читайте также: Клеевая ткань в авто
Гексокиназа отличается от глюкокиназы высоким сродством к глюкозе. Следовательно, этот фермент, в отличие от глюкокиназы, активен при низкой концентрации глюкозы в крови, что характерно для постабсорбтивного состояния. Печень в этот период поглощает гораздо меньше глюкозы, так как скорость её внутриклеточного фосфорилирования глюкокиназой резко снижается. Тогда как потребление глюкозы мозгом, эритроцитами и другими тканями обеспечивается активной в этих условиях гексокиназой. Фермент гексокиназа может катализировать фосфорилирование не только D-глюкозы, но и других гексоз, хотя и с меньшей скоростью. Активность гексокиназы изменяется в зависимости от потребностей клетки в энергии. В качестве регуляторов выступают соотношение АТФ/АДФ и внутриклеточный уровень глюкозо-6-фосфата (продукта катализируемой реакции). При снижении расхода энергии в клетке повышается уровень АТФ (относительно АДФ) и глюкозо-6-фосфата. В этом случае активность гексокиназы снижается, и, следовательно, уменьшается скорость поступления глюкозы в клетку. Следует отметить, что в разных тканях гексокиназа присутствует в различных изоформах, отличающихся величиной Кm. Глюкокиназа печени (и почек) является изоформой IV (гексокиназа IV). В клетках мышц содержится гексокиназа II, а в клетках опухолевых тканей преобладает гексокиназа III, с более высоким, чем у гексокиназы II, сродством к глюкозе.
Дефосфорилирование глюкозо-6-фосфата Превращение глюкозо-6-фосфата в глюкозу возможно в печени, почках и клетках эпителия кишечника. В клетках этих органов имеется фермент глюкозо-6-фосфатаза, катализирующая отщепление фосфатной группы гидролитическим путём: Глюкозо-6-фосфат +Н2О → Глюкоза + Н3РО4
Образовавшаяся свободная глюкоза способна диффундировать из этих органов в кровь. В других органах и тканях глюкозо-6-фосфатазы нет, и поэтому дефосфорилирование глюкозо-6-фосфата невозможно. Пример подобного необратимого проникновения глюкозы в клетку — мышцы, где глюкозо-6-фосфат может использоваться только в метаболизме этой клетки.
Метаболизм глюкозо-6-фосфата Глюкозо-6-фосфат может использоваться в клетке в различных превращениях, основными из которых являются: синтез гликогена, катаболизм с образованием СО2 и Н2О или лактата, синтез пентоз. Распад глюкозы до конечных продуктов служит источником энергии для организма. Вместе с тем в процессе метаболизма глюкозо-6-фосфата образуются промежуточные продукты, используемые в дальнейшем для синтеза аминокислот, нуклеотидов, глицерина и жирных кислот. Таким образом, глюкозо-6-фосфат — не только субстрат для окисления, но и строительный материал для синтеза новых соединений
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом