
 Обмен белков в тканях.
Обмен белков в тканях.
Основная часть аминокислот, которые образуются в кишечнике из белков, поступает в кровь (95%) и небольшая часть — в лимфу. По воротной вене аминокислоты попадают в печень, где расходуются на биосинтез различных специфических белков (альбуминов, глобулинов, фибриногена). Другие аминокислоты током крови разносятся ко всем органам и тканям, транспортируются внутрь клеток, где они используются для биосинтеза белков.
Неиспользованные аминокислоты окисляются до конечных продуктов обмена. Процесс расщепления тканевых белков катализируется тканевыми ферментами – протеиназами — катепсинами (часто их называют тканевыми протеазами).
Соотношение между аминокислотами в белках, которые распадаются и синтезируются, разное, поэтому часть свободных аминокислот должна быть преобразована в другие аминокислоты или окислена до простых соединений и выведена из организма.
Итак, в организме существует внутриклеточный запас аминокислот, которые в значительной мере пополняется за счет процессов взаимопревращения аминокислот, гидролиза белков, синтеза аминокислот и поступления их из внеклеточной жидкости. В то же время благодаря синтезу белков и другим реакцям (образование мочевины, пуринов и т.п.) постоянно происходит удаление свободных аминокислот из внеклеточной жидкости.
Пути обмена аминокислот в тканях.
В основе различных путей обмена аминокислот лежат три типа реакций: по аминной и карбоксильной группам и по боковой цепи. Реакции по аминной группе включают процессы дезаминирования, переаминирования, аминирования , по карбоксильной группе — декарбоксилирование. Безазотистая часть углеродного скелета аминокислот подвергается различным превращениям с образованием соединений, которые затем могут включаться в цикл Кребса для дальнейшего окисления.
Пути внутриклеточного превращения аминокислот сложны и перекрещиваются со многими другими реакциями обмена, в результате чего промежуточные продукты обмена аминокислот могут служить необходимыми предшественниками для синтеза различных компонентов клеток и быть биологически активными веществами.
Катаболизм аминокислот у млекопитающих (и у человека) происходит, в основном, в печени и немного слабее в почках.
Дезаминирование аминокислот.
Суть дезаминирования заключается в расщеплении аминокислот под действием ферментов на аммиак и безазотистый остаток (жирные кислоты, оксикислоты, кетокислоты). Дезаминирование может идти в виде восстановительного, гидролитического, окислительного и внутримолекулярного процессов. Последние два типа превалируют у человека и животных.
Окислительное дезаминирование подразделяется на две стадии. Первая стадия является ферментативной, она заканчивается образованием неустойчивого промежуточного продукта – иминокислоты (карбоновые кислоты, содержащие иминогруппу (=NH), которая во второй стадии спонтанно в присутствии воды распадается на аммиак и aльфа-кетокислоту. Ферменты, которые катализируют этот процесс, содержат в качестве простетической группы (органические соединение небелковой природы) НАД (никотинамидадениндинуклеотид) или ФАД (флавинадениндинуклеотид).
В организме человека наиболее активно протекает дезаминирование глутаминовой кислоты под действием фермента глутаматдегидрогеназы , которая находится в митохондриях клеток всех тканей. В результате этого процесса образует альфа-кетоглутаровая кислота, которая участвует во многих процессах обмена веществ.
Трансаминирование (переаминирование) аминокислот.
Обязательным условием трансаминирования является участие дикарбоновых аминокислот (глутаминовой и аспарагиновой), которые в виде соответствующих им кетокислот — альфа-кетоглутаровой и щавелевоуксусной могут взаимодействовать со всеми аминокислотами, за исключением лизина, треонина и аргинина.
При переаминировании происходит непосредственный перенос аминогруппы с аминокислоты на кетокислоту, а кетогруппы — с кетокислоты на аминокислоту без освобождения при этом аммиака. Этот процесс протекает в несколько этапов. Реакцию катализируют ферменты, относящиеся к классу трансфераз, их простетической группой является фосфорпиридоксаль-фосфорный эфир витамина В6. Процесс переаминирования широко распространен в живой природе. Его особенность — легкая обратимость.
Реакции переаминирования играют большую роль в обмене веществ. От них зависят такие важнейшие процессы, как биосинтез многих заменимых аминокислот из соответствующих им кетокислот, распад аминокислот, объединение путей углеводного и аминокислотного обмена, когда из продуктов распада глюкозы, например, пировиноградной кислоты, может образоваться аминокислота аланин и наоборот.
Восстановительное аминирование.
Этот процесс противоположен дезаминированию. Он обеспечивает связывание аммиака кетокислотами с образованием соответствующих аминокислот. Восстановительное аминирование катализируется хорошо функционирующей ферментной системой, обеспечивающей аминирование aльфа-кетоглутаровой или щавелевоуксусной кислоты с образованием глутаминовой или аспарагиновой кислоты.
Читайте также: Какие ткани подходят для штор в гостиную
При обезвреживании аммиака неорганическими и органическими кислотами происходит образование аммонийных солей. Этот процесс осуществляется в почках. Образовавшиеся аммонийные соли выводятся из организма с мочой и потом.
Декарбоксилирование аминокислот.
Процесс декарбоксилирования катализируется декарбоксилазами, специфическими для каждой аминокислоты, простетической группой которых служит пиридоксальфосфат. Эти ферменты относятся к классу лиаз. Процесс декарбоксилирования, который заключается в отщеплении от аминокислот СО2 с образованием аминов, можно показать на следующей схеме:

Механизм реакции декарбоксилирования аминокислот согласно общей теории пиридоксалевого катализа сводится к образованию пиридоксальфосфат-субстратного комплекса в активном центре фермента.
Таким путем из триптофана образуется триптамин, из гидрокситриптофана — серотонин. Из аминокислоты гистидина образуется гистамин . Из глутаминовой кислоты при декарбоксилировании образуется гамма-аминомасляная кислота (ГАМК) .
Амины, образованные из аминокислот, называют биогенными аминами, так как они оказывают на организм мощный биологический эффект. Биогенные амины проявляют физиологическое действие в очень малых концентрациях. Так, введение в организм гистамина приводит к расширению капилляров и повышению их проницаемости, сужению крупных сосудов, сокращению гладких мышц различных органов и тканей, повышению секреции соляной кислоты в желудке. Кроме того, гистамин участвует в передаче нервного возбуждения.
Серотонин способствует повышению кровяного давления и сужению бронхов; его малые дозы подавляют активность центральной нервной системы, в больших дозах это вещество оказывает стимулирующее действие. В различных тканях организма большие количества гистамина и серотонина находятся в связанной, неактивной форме. Биологическое действие они проявляют только в свободной форме.
Гамма-аминомасляная кислота (ГАМК) накапливается в мозговой ткани и представляет собой нейрогуморальный ингибитор-медиатор торможения центральной нервной системы.
Большие концентрации этих соединений могут представлять угрозу для нормального функционирования организма. Однако в животных тканях имеется аминоксидаза , расщепляющая амины до соответствующих альдегидов, которые потом превращаются в жирные кислоты и распадаются до конечных продуктов.
«Обмен аминокислот в тканях» — это третья статья из цикла «Обмен белков в организме человека». Первая статья – « Расщепление белков в пищеварительном тракте ». Вторая статья « Обезвреживание продуктов гниения белков в кишечнике ».
2 Трансаминирование аминокислот
Трансаминирование аминокислот — основной путь дезаминирования аминокислот, который происходит без образования свободного NH3. Это обратимый процесс переноса NH2–группы с аминокислоты на –кетокислоту. Процесс открыли А.Е. Браунштейн и М.Б. Крицман (1937).
В трансаминировании могут принимать участие все аминокислоты, кроме треонина, лизина, пролина и гидроксипролина.
Реакция трансаминирования в общем виде выглядит следующим образом:







 СООН СООН СООН СООН
СООН СООН СООН СООН



 НС — NH2 + C = O C = O + НС — NH2
НС — NH2 + C = O C = O + НС — NH2
Ферменты, которые катализируют реакции этого типа, называются аминотрансферазами (трансаминаза–ми). В организме человека функционируют аминотрансфе–разы L–аминокислот. Акцептором аминогруппы в реакции являются -кетокислоты – пируват, оксалоацетат, -кето–глутарат. Наиболее распространенные аминотрансферазы – АлАТ (аланинаминотрансфераза), АсАТ (аспартатамино–трансфераза), тирозинаминотрансфераза.
Реакция, которую катализирует фермент АлАТ, представлена ниже:
Реакцию, которую катализирует фермент АсАТ, схематически можно изобразить следующим образом:
Кофермент трансаминаз – пиридоксальфосфат (В6) – входит в состав активного центра фермента. В процессе трансаминирования кофермент выполняет роль перенос–чика аминогруппы, и происходит взаимопревращение двух коферментных форм ПАЛФ(пиридоксаль–5–ф) и ПАМФ (пиридоксамин–5–ф):
Палф памф.
Трансаминирование активно протекает в печени. Это позволяет регулировать концентрацию любых амино–кислот в крови, в том числе и поступивших с пищей (за исключением тре, лиз, про). Благодаря этому оптимальная смесь аминокислот переносится с кровью во все органы.
Некоторые клинические аспекты
В ряде случаев может происходить нарушение трансаминирования аминокислот:
2) при лечении туберкулеза антагонистами трансами–аз – фтивазидом и его аналогами;
3) при голодании, циррозе и стеатозе печени наблюда–ется недостаток синтеза белковой части трансами–наз.
Для диагностики имеет значение определение активности аминотрансфераз в плазме крови. При патологических состояниях происходит усиление цитолиза в том или ином органе, что сопровождается повышением активности этих ферментов в крови.
Читайте также: Как соединить ткани для простыни
Отдельные трансаминазы находятся в различных тканях в неодинаковом количестве. АсАТ больше в кардиомиоцитах, печени, скелетных мышцах, почках, поджелудочной железе. АлАТ – в рекордном количестве в печени, в меньшей степени — в поджелудочной железе, миокарде, скелетной мускулатуре. Следовательно, повышение активности АсАТ в крови более характерно для инфаркта миокарда (ИМ), а повышение активности АлАТ может свидетельствовать о цитолизе в гепатоцитах. Так, при остром инфекционном гепатите в крови активность АлАТ >АсАТ; но при циррозе печени -АсАТ >АлАТ. Незначительное повышение активности АлАТ имеет место также при ИМ. Поэтому определение активности сразу двух трансаминаз является важным диаг–ностическим тестом. В норме соотношение активностей АсАТ/АлАТ (коэффициент де Ритиса) составляет 1,330,42. При ИМ величина этого коэффициента резко возрастает, у больных инфекционным гепатитом, напротив, происходит снижение этого показателя.
Пути обмена аминокислот в тканях
Пути обмена аминокислот в тканях
Аминокислоты – это бифункциональные соединения, содержащие аминную и карбоксильную группу. Реакции по этим группам являются общими для различных аминокислот.
К ним относят:
1. по аминной группе – реакции дезаминирования и трансаминирования;
2. по карбоксильной группе – реакции декарбоксилирования.
Кроме этих общих путей возможны реакции по углеводородному радикалу аминокислот, которые являются специфическими для каждой аминокислоты.
Катаболизм большинства аминокислот начинается с отщепления a-аминогруппы, которое возможно в реакциях трансаминировани и дезаминирования.
Трансаминирование – реакции переноса a-аминогруппы с аминокислоты на a-кетокислоту, в результате чего образуются новая кетокислота и новая аминонокислота. Реакции катализируют ферменты аминотрансферазы. Это сложные ферменты, коферментом которых является производное витамина В 6 – пиридоксальфосфат, который обратимо может переходить в пиридоксаминфосфат. Реакции трансаминирования обратимы, и могут проходить как в цитоплазме, так и в митохондриях клеток. В клетках человека найдено более 10 аминотрансфераз, отличающихся по субстратной специфичности. Вступать в реакции трансаминирования могут почти все аминокислоты, за исключением лизина, треонина и пролина.
Реакции трансаминирования протекают в 2 стадии. На первой стадии к пиридоксальфосфату в активном центре фермента присоединяется аминогруппа от первого субстрата – аминокислоты. Образуется комплекс фермент- пиридоксаминфосфат и кетокислота – первый продукт реакции. Этот процесс включает промежуточное образование 2 шиффовых оснований (альдимин и кетимин).
На второй стадии пиридоксаминфосфат соединяется с новой кетокислотой (второй субстрат) и снова через промежуточное образование 2 шиффовых оснований передает аминогруппу на кетокислоту. В результате фермент возвращается в свою нативную форму, и образуется новая аминокислота – второй продукт реакции.
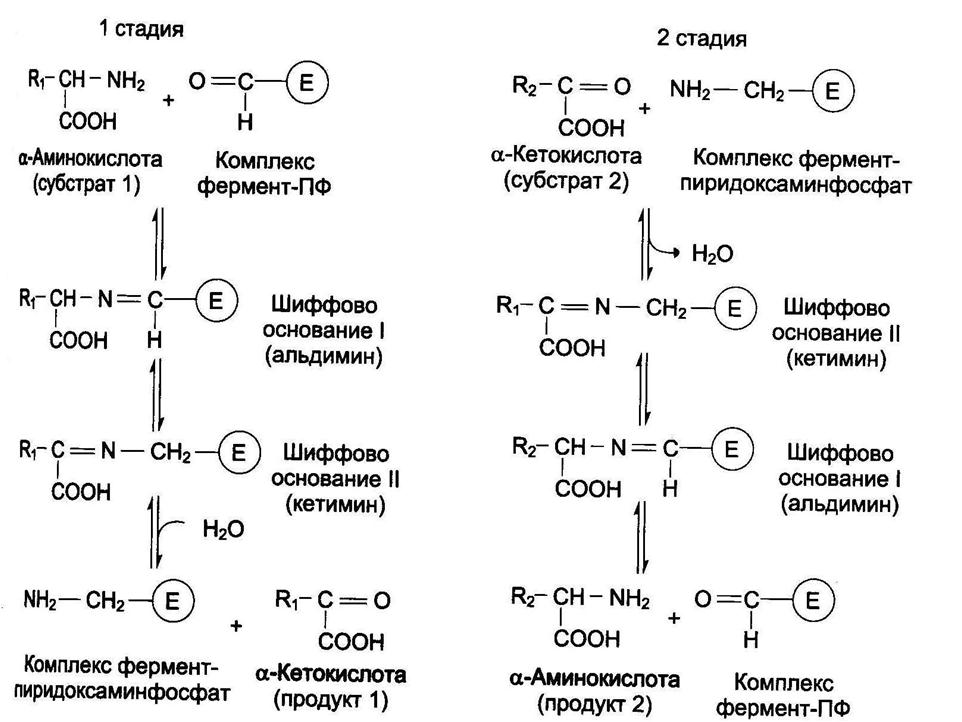
Чаще всего в реакциях трансаминирования участвуют аминокислоты, содержание которых в тканях значительно выше остальных – глутамат, аланин, аспартат. Наиболее распространенными в большинстве тканей являются аланинаминотрансфераза (АлАТ) и аспартатаминотрансфераза (АсАТ).
Наибольшая активность АсАТ обнаруживается в клетках сердечной мышцы и печени, в то время как в крови обнаруживается только фоновая активность АлАТ и АсАТ. Поэтому можно говорить об органоспецифичности этих ферментов, что позволяет их широко примененятьих с диагностической целью (при инфарктах миокарда и гепатитах).
Биологическое значение трансаминирования
Трансаминирование – первая стадия дезаминирования большинства аминокислот, т.е. начальный этап их катаболизма. Образующиеся при этом кетокислоты окисляются в ЦТК или используются для синтеза глюкозы и кетоновых тел. Поскольку этот процесс обратим, ферменты аминотрансферазы функционируют как в процессах катаболизма, так и биосинтеза аминокислот. Трансаминирование – заключительный этап синтеза заменимых аминокислот из соответствующих кетокислот, если они необходимы в данный момент клеткам. В результате происходит перераспределение аминнного азота в тканях. При трансаминированиии общее количество аминокислот в клетке не меняется.
При физиологических значениях рН в тканях высоко активны оксидазы D-аминокислот. Они также обнаружены в почках и печени и находятся в микросомах. Роль оксидаз D-аминокислот невелика и до конца не понятна, потому что в белки пищи и тканей человека входят только природные L-аминокислоты.
Читайте также: Методы определения волокнистого состава тканей кратко
В печени человека присутствуют специфические ферменты, катализирующие реакции дезаминирования серина, треонина, цистеина и гистидина неокислительным путем.
Данный текст является ознакомительным фрагментом.
Читайте также
Глава 496. Почему кодируемых аминокислот двадцать? (XII)
Глава 496. Почему кодируемых аминокислот двадцать? (XII) Неискушенному Читателю может показаться, что элементы машины генетического кодирования описаны в предыдущей главе настолько детально, что к концу чтения он стал даже как-то утомляться, чувствуя, что несколько
Всасывание аминокислот.
Всасывание аминокислот. Происходит путем активного транспорта с участием переносчиков. Максимальная концентрация аминокислот в крови достигается через 30–50 мин после приема белковой пищи. Перенос через щеточную каемку осуществляется целым рядом переносчиков, многие
Наследственные нарушения транспорта аминокислот
Наследственные нарушения транспорта аминокислот Болезнь Хартнупа – нарушение всасывания триптофана в кишечнике и его реабсорбции в почечных канальцах. Так как триптофан служит исходным продуктом для синтеза витамина РР, то основные проявления болезни Хартнупа –
Расщепление белков в тканях
Расщепление белков в тканях Осуществляется с помощью протеолитических лизосомальных ферментов катепсинов. По строению активного центра выделяют цистеиновые, сериновые, карбоксильные и металлопротеиновые катепсины. Роль катепсинов:1. создание биологически активных
Превращение аминокислот микрофлорой кишечника
Превращение аминокислот микрофлорой кишечника Микроорганизмы кишечника располагают набором ферментативных систем, отличных от соответствующих ферментов тканей организма человека и катализирующих самые разнообразные превращения пищевых аминокислот и не
Трансаминирование аминокислот
Трансаминирование аминокислот Трансаминирование – реакции переноса a-аминогруппы с аминокислоты на a-кетокислоту, в результате чего образуются новая кетокислота и новая аминонокислота. Реакции катализируют ферменты аминотрансферазы. Это сложные ферменты, коферментом
Дезаминирование аминокислот
Дезаминирование аминокислот Дезаминирование аминокислот – реакция отщепления a-аминогруппы от аминокислоты с выделением аммиака. Различают два типа реакций дезаминирования: прямое и непрямое.Прямое дезаминирование – непосредственное отщепление аминогруппы от
Непрямое дезаминирование аминокислот
Непрямое дезаминирование аминокислот Большинство аминокислот не способно дезаминироваться в одну стадию, подобно глутамату. Аминогруппы таких аминокислот перносятся на ?-кетоглутарат с образованием глутаминовой кислоты, которая затем подвергается прямому
Декарбоксилирование аминокислот
Декарбоксилирование аминокислот Некоторые аминокислоты и их производные могут подвергаться декарбоксилированию. Реакции декарбоксилирования необратимы и катализируются ферментами декарбоксилазами, нуждающимися в пиридоксальфосфате в качестве кофермента.
Пути катаболизма углеродного скелета аминокислот
Пути катаболизма углеродного скелета аминокислот Трансаминирование и дезаминирование аминокислот ведет к образованию безазотистых углеродных скелетов аминокислот – ?-кетокислот. В состав белков входят 20 аминокислот, различающихся по строению углеводородного
Глава 25. Метаболизм отдельных аминокислот
Глава 25. Метаболизм отдельных аминокислот Метаболизм метионина Метионин – незаменимая аминокислота. Метильная группа метионина – мобильный одноуглеродный фрагмент, используемый для синтеза ряда соединений. Перенос метильной группы метионина на соответствующий
Распад нуклеиновых кислот в желудочно-кишечном тракте и тканях
Распад нуклеиновых кислот в желудочно-кишечном тракте и тканях Нуклеиновые кислоты поступают в организм с пищей главным образом в составе нуклеопротеинов и высвобождаются в результате действия протеолитических ферментов желудочно-кишечного тракта. Далее под
Роль печени в обмене аминокислот и белков
Роль печени в обмене аминокислот и белков Печень играет центральную роль в обмене белков и других азотсодержащих соединений. Она выполняет следующие функции:1. синтез специфических белков плазмы: — в печени синтезируется: 100 % альбуминов, 75 – 90 % ?-глобулинов, 50 %
Обмен свободных аминокислот в головном мозге
Обмен свободных аминокислот в головном мозге Аминокислоты играют важную роль в метаболизме и функционировании ЦНС. Это объясняется не только исключительной ролью аминокислот как источников синтеза большого числа биологически важных соединений, таких как белки,
Глава 496. Почему кодируемых аминокислот двадцать? (XII)
Глава 496. Почему кодируемых аминокислот двадцать? (XII) Неискушенному Читателю может показаться, что элементы машины генетического кодирования описаны в предыдущей главе настолько детально, что к концу чтения он стал даже как-то утомляться, чувствуя, что несколько
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности