
С тех пор процессы пролиферации и дифференцировки клеток ушли далеко вперед, создав настоящее чудо — вас, человека. У растения жизнь начинается точно так же — с одной маленькой клетки, из которой в дальнейшем будут развиваться ткани и органы самых разных форм. Главная заслуга роста растения принадлежит образовательной ткани.
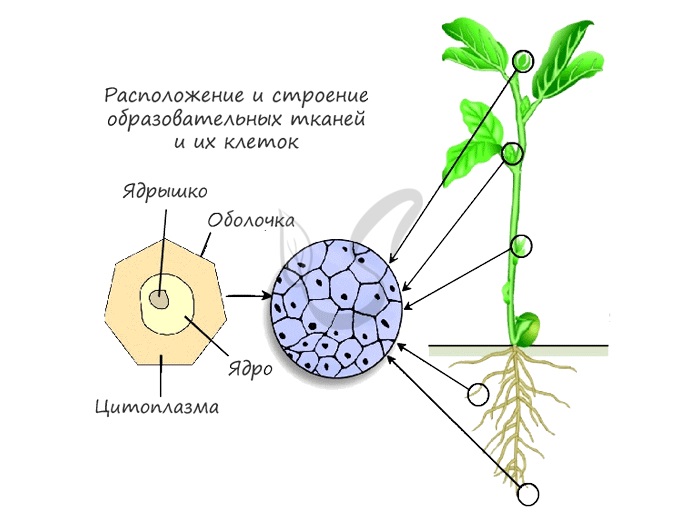
Как вы видите, на картинке схематично изображено месторасположение образовательной ткани. Главным образом это:
- Кончик побега — конус нарастания в почках
- Кончик корня — зона деления, прикрытая корневым чехликом для защиты
- Камбий — обеспечивает рост растения в ширину
- Основание междоузлий и черешков листьев — это также зоны активного роста растения
Именно в этих местах и происходит деление клеток и рост растения. Важно отметить, что сезонные изменения активности клеток камбия являются причиной возникновения годичных колец древесины. Внешний вид годичных колец обусловлен хронологической закономерностью: весной больше образуется проводящей ткани (более тонкая и рыхлая внутри), а осенью — механическая (толстая, более твердая). Именно поэтому годичные кольца на спиле дерева выглядят как чередование колец, отличающихся друг от друга.
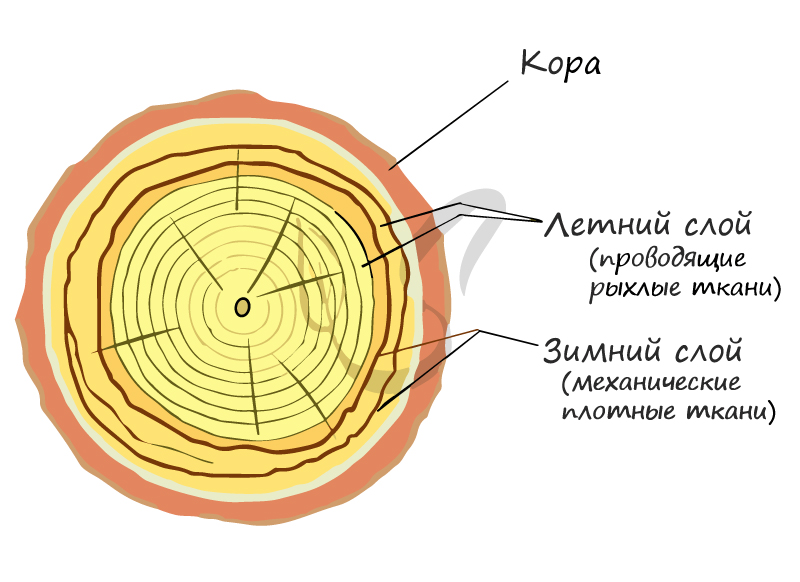
На внешний вид годичных колец оказывают весьма сильное влияние условия внешней среды. Так, при дефиците трофического компонента (питательных веществ), к примеру, у растений, растущих на болоте, годичные кольца выглядят тоньше своих обычных размеров.
Ветер также оказывает существенное влияние: при его постоянном действии происходит перераспределение древесины по стволу. Оказывая действие на крону, ветер смещает центр тяжести дерева, что сказывается на его нижележащих отделах. Они начинают компенсаторно утолщаться для предотвращения слома дерева. При постоянно дующем ветре ствол сильно искривляется, а форма кроны становится флагообразной.

Тема камбия и форм стволов растений весьма занимательна, и все-таки мы должны разобраться в строении самой образовательной ткани. Она представлена живыми мелкими быстро делящимися клетками с относительно крупным ядром. Объем цитоплазмы небольшой, она вязкая по консистенции, оболочка клетки тонкая. Это уязвимые клетки, которые растение оберегает по-своему, подобно тому, как животные оберегают только что появившееся потомство.

Другое название образовательных тканей — меристемы (с др.-греч. — «μεριστός» — делимый). По времени возникновения различают первичные и вторичные меристемы.
Первичные меристемы — закладываются в эмбриогенезе
1) Вставочные меристемы (интеркалярные) — в виде отдельных участков в зоне активного роста в разных частях растения. Такие ткани можно найти в основании междоузлий у злаков, черешков листьев у многих растений. У злаковых наблюдается быстрый рост стебля за счет множественного расположения данной ткани на стебле — «вставочный рост».
2) Прокамбий — основа будущего камбия, перицикла, окружающего проводящие ткани в один или несколько слоёв (у голосеменных). В корнях перицикл является корнеродным слоем, так как в корне с него начинается формирование осевого цилиндра, наружным слоем которого он является. В нём закладываются придаточные и боковые корни, что имеет принципиальное значение для формирования корневой системы растения.
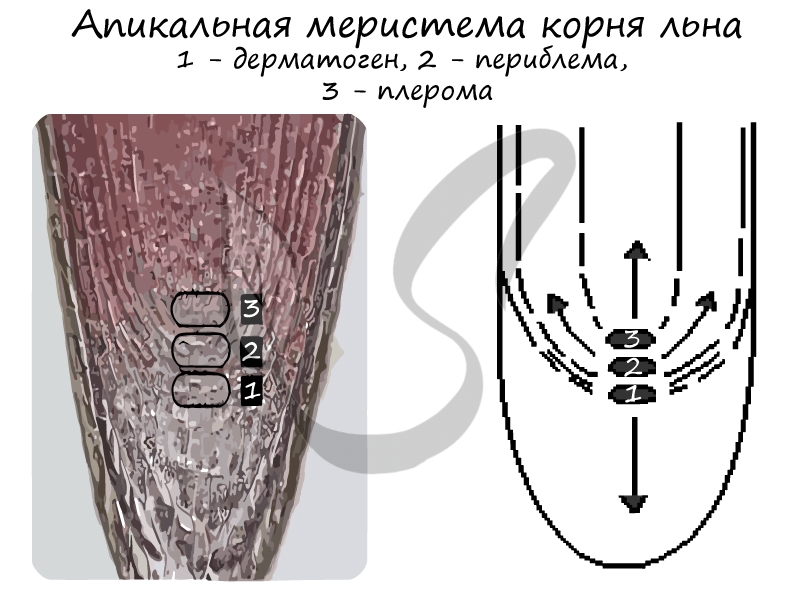
3) Верхушечные (апикальные) — формируются на верхушках стеблей и кончиках корней. В периферической части корня различают три слоя:
- Дерматоген — в дальнейшем преобразующийся в первичную покровно-всасывающую ризодерму (эпиблему или ризодерму)
- Периблема — образующая ткани первичной коры
- Плерома — внутренний слой ткани центрального осевого цилиндра
Вторичные меристемы — закладываются в постэмбриональном развитии
Камбий и феллоген (пробковый камбий) — занимают боковое положение по отношению к оси органа, обеспечивают рост вширь. Растения часто повреждаются, их задевают животные, нарушая целостность тканей и органов. На этот случай в группе вторичных меристем есть раневые меристемы, дающие начало защитной ткани в местах повреждения растения.

Топографическая классификация меристем
Спешу заверить, это отнюдь не сложная классификация, которой нужно бояться. Речь пойдет о взгляде на те же образовательные ткани с другой стороны. В переводе с греч. τόπος — место. Мы рассмотрим меристемы в соответствии с их месторасположением на растении.
- Верхушечная или апикальная (лат. apex — вершина) — расположена на кончике корня и конусе нарастания побега
- Боковая или латеральная (лат. latus — бок): камбий – обеспечивает рост стебля и корня в толщину
- Краевая или маргинальная (лат. margo — край) меристема даёт начало листовой пластинке
- Вставочная или интеркалярная (лат. inter — между и calaris — вставочный, добавочный) — расположена преимущественно у основания стеблевых междоузлий между зонами дифференцированных тканей.
Читайте также: Шенилловая ткань что это
© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Раневые ткани вторичные или первичные
Существуют три классических типа заживления раны: заживление первичным натяжением; вторичным заживлением и заживление под струпом.
Заживление раны первичным натяжением характеризуется сращением краев раны путем соединительнотканной организации грануляционной ткани, которая прочно соединяет стенки раны. Рубец после заживления раны первичным натяжением ровный, гладкий, почти незаметный.
Заживление первичным натяжением возможно при плотном соприкосновении краев раны, сохранении их жизнеспособности и, главное, асептичности раны. Первичным натяжением заживают послеоперационные раны или небольшие резаные раны, когда края отстоят друг от друга не больше чем на 1 см.

Заживление раны вторичным натяжением происходит при обширных ранах, наличии в ране нежизнеспособных тканей, развитии инфекции. В отдельных участках дна раны появляются островки: грануляций, которые постепенно заполняют всю рану и начинают реорганизовываться в рубец. Параллельно идёт процесс эпители-зации от краёв раны.
Заживление раны вторичным натяжением всегда заканчивается формированием более или менее выраженного рубца. Чем выра-женнее была инфекция, тем более грубым будет рубец.
Заживление под струпом происходит обычно при незначительных ссадинах, потёртостях, небольших ожогах I—II степени. Струп образуется в результате свёртывания излившейся крови и лимфы, под ним происходит быстрая регенерация эпидермиса, после чего струп отторгается. Если не присоединилась инфекция, то после заживления раны под струпом следа не остаётся.
Таким образом, заживление раны определяется наличием или отсутствием инфекции в ране. Степень инфицированности раны в свою очередь зависит от присутствия в ране некротически изменённых тканей.
Первичные меристемы
Первичные меристемы происходят непосредственно из меристемы зародыша и обладают способностью к делению. По положению в растении первичные меристемы могут быть верхушечными (апикальными) (рис. 2.2), вставочными (интеркалярными) и боковыми (латеральными).
Верхушечные (апикальные) меристемы — такие меристемы, которые располагаются у взрослых растений на верхушках стеблей и кончиках корней и обеспечивают рост тела в длину. У стеблей в конусе нарастания выделяют два меристематических слоя: тунику, из которой образуются покровная ткань и периферическая часть первичной коры, и корпус, из которого образуются внутренняя часть первичной коры и центральный осевой цилиндр (рис. 2.3).
В кончике корня различают три слоя:
1) дерматоген, из которого образуется первичная покровно-всасывающая ткань — ризодерма;
2) периблему, из которой развиваются ткани первичной коры;
3) плером, образующий ткани центрального осевого цилиндра (рис. 2.4).
Рис. 2.2. Верхушечная меристема побега: 1 — конус растения; 2 — зачатки листьев; 3 — зачаток боковой почки
Боковые (латеральные) меристемы по происхождению могут быть первичными и вторичными, на поперечном срезе осевых органов имеют вид колец. Примером первичной боковой меристемы служат прокамбий и перицикл. Из прокамбия формируются камбий и первичные элементы сосудисто-волокнистых пучков (первичная флоэма и первичная ксилема), при этом клетки прокамбия непосредственно дифференцируются в клетки первичных проводящих тканей.
Боковые меристемы располагаются параллельно поверхности органа и обеспечивают рост осевых органов в толщину.
Читайте также: Костная ткань развивается из мезенхимы в участках лишенных кровеносных сосудов
Вставочные (интеркалярные) меристемы чаще первичные и сохраняются в виде отдельных участков в зонах активного роста в различных частях растения (например, в основании черешков листьев, у оснований междоузлий). В основании междоузлий у злаков деятельность этой меристемы ведет к удлинению междоузлий, что обеспечивает рост стебля в длину.
Вторичные меристемы
К вторичным относят боковые и раневые меристемы.
Боковые (латеральные) меристемы представлены камбием и феллогеном. Они формируются из промеристем (прокамбия) или постоянных тканей путем их дедифференцировки. Клетки камбия делятся перегородками, параллельными поверхности органа (периклинально). Из клеток, отложенных камбием наружу, развиваются элементы вторичной флоэмы, а из клеток, отложенных внутрь, — вторичной ксилемы. Камбий, возникший из постоянных тканей путем дедифференцировки, называют добавочньм. По строению и функции он не отличается от камбия, возникшего из промеристем. Феллоген формируется из постоянных тканей, расположенных в субэпидермальных слоях (под эпидермой). Делясь периклинально, феллоген отделяет наружу будущие клетки пробки (феллемы), а внутрь — клетки феллодермы. Таким образом, феллоген формирует вторичную покровную ткань — перидерму. Боковые меристемы располагаются параллельно поверхности органа и обеспечивают рост осевых органов в толщину.
Раневые меристемы образуются при повреждении тканей и органов. Вокруг повреждения живые клетки дедифференцируются, начинают делиться и тем самым превращаются во вторичную меристему. Их задача — образовать плотную защитную ткань, состоящую из паренхимных клеток, — каллюс. Эта ткань беловатого или желтоватого цвета, ее клетки имеют крупные ядра и достаточно толстые клеточные стенки. Каллюс возникает при прививках, обеспечивая срастание привоя с подвоем, и в основании черенков. В нем могут закладываться придаточные корни и почки, поэтому его используют для получения культуры изолированных тканей.
На поверхности всех органов растений находятся покровные ткани. Основная их функция — защитная. Они защищают внутренние ткани от неблагоприятных факторов: механических воздействий, резкой смены температур, проникновения вредителей и патогенных микроорганизмов, излишнего испарения и иссушения. Кроме защитной, покровные ткани выполняют и другие функции — газообмена, транспирации, выделения, поглощения и др.
Различают первичную, вторичную и третичную покровные ткани.
Первичная покровная ткань
Она покрывает листья, цветки, плоды, а также стебли при их первичном строении.
Эпидерма выполняет в основном функции покровных тканей. С полифункциональностью связано разнообразие ее структур, т. е. кожица — сложная ткань. Она состоит из следующих компонентов:
1. Основная ткань, клетки которой образуют большую часть ее поверхности.
2. Устьица и связанные с ними побочные клетки.
3. Различные выросты (волоски, трихомы).
К первичной покровной ткани относят эпидерму собственно эпидермальных, околоустьичных, замыкающих клеток устьица и трихом.
Клетки основной ткани кожицы паренхимные, плотно сомкнутые, иногда имеют извилистые стенки, что способствует более прочному их соединению. Эпидерма органов, вытянутых в длину (стебли, черешки, ланцетовидные листья), состоит из клеток, удлиненных параллельно продольной оси органа. Боковые и внутренняя стенки клеток тонкие целлюлозные.
Наружные стенки клеток эпидермы покрыты кутикулой. Она защищает клетки от механических повреждений, проникновения паразитов и поглощает ультрафиолетовые лучи. Кутинизация рассматривается как приспособление для уменьшения испарения и вымывания веществ из клеток выпадающими осадками.
Пектиновые вещества и целлюлоза, входящие в клеточную стенку, могут подвергаться ослизнению с образованием слизей и камедей. Они представляют собой полимерные углеводы, родственные пектиновым веществам, и отличаются способностью к сильному набуханию при соприкосновении с водой. Камеди в набухшем состоянии клейкие и могут вытягиваться в нити, тогда как слизи сильно расплываются и в нити не вытягиваются. Пектиновые слизи встречаются у представителей семейств лилейных, крестоцветных, мальвовых, липовых, розоцветных, в отличие от целлюлозных слизей, встречающихся гораздо реже (например, у орхидных).
Газообмен между внутренними тканями растений и внешней средой, а также испарение воды в эпидерме осуществляются через специальные образования — устьица.
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток бобовидной формы и устьичной щели (своеобразного межклетника между ними). Имеются главным образом в листьях, но встречаются и в стебле (рис. 2.6).
Читайте также: Постельные ткани для отелей турции
Устьица расположены на нижней стороне листа, но у водных растений с плавающими листьями они находятся только на верхней стороне листа. По форме клеток эпидермы листа и расположению устьиц можно отличить однодольное растение от двудольного (рис. 2.8). Собственно эпидермальные клетки листа двудольных растений в очертаниях волнистые (рис. 2.9), а у однодольных они вытянутые, ромбической формы.
Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью расположена крупная дыхательная полость (межклетник), окруженная клетками мезофилла листа.
Замыкающие клетки окружены околоустьичными клетками, которые совместно образуют устьичный комплекс (рис. 2.7). Различают следующие основные типы устьичных комплексов:
1) аномоцитный (беспорядочный) — замыкающие клетки не имеют ярко выраженных околоустьичных клеток; характерен для всех высших растений, исключая хвойные;
2) анизоцитный (неравноклеточный) — замыкающие клетки устьица окружены тремя околоустьичными клетками, одна из которых намного крупнее (или меньше) остальных;
3) парацитный (параллельно-клеточный) — одна околоустьичная клетка (или более) расположена параллельно замыкающим;
4) диацитный (перекрестно-клеточный) — две околоустьичные клетки расположены перпендикулярно замыкающим;
5) тетрацитный (от греч. tetra — четыре) — главным образом у однодольных;
Рис. 2.7. Основные типы устьичного аппарата: 1 — аномоцитный (у всех высших растений, кроме хвощей); 2 — диацитный (у папоротников и цветковых); 3 — парацитный (у папоротников, хвощей, цветковых и гнетовых); 4 — анизоцитный (только у цветковых); 5 — тетрацитный (главным образом у однодольных); 6 — анциклоцитный (у папоротников, голосеменных и цветковых)
Механизм работы устьиц обусловлен осмотическими свойствами клеток. При освещении поверхности листа солнцем в хлоропластах замыкающих клеток происходит активный процесс фотосинтеза. Насыщение клеток продуктами фотосинтеза, сахарами влечет за собой активное поступление в клетки ионов калия, вследствие чего концентрация клеточного сока в замыкающих клетках возрастает. Возникает разность концентрации клеточного сока околоустьичных и замыкающих клеток. В силу осмотических свойств клеток вода из околоустьичных клеток поступает в замыкающие, что ведет к увеличению объема последних и резкому возрастанию тургора. Утолщение «брюшных» стенок замыкающих клеток, обращенных к устьичной щели, обеспечивает неравномерное растяжение клеточной стенки; замыкающие клетки приобретают выраженную бобовидную форму, и устьичная щель открывается. При снижении интенсивности фотосинтеза (например, вечером) уменьшается образование сахаров в замыкающих клетках. Приток ионов калия прекращается. Концентрация клеточного сока в замыкающих клетках снижается по сравнению с околоустьичными. Вода путем осмоса уходит из замыкающих клеток, понижая их тургор, в результате ночью устьичная щель закрывается.
Клетки эпидермы плотно сомкнуты между собой, благодаря этому эпидерма выполняет целый ряд функций:
• препятствует проникновению болезнетворных организмов внутрь растения;
• защищает внутренние ткани от механических повреждений;
• регулирует газообмен и транспирацию;
• через нее выделяются вода, соли;
• может функционировать как всасывающая ткань;
принимает участие в синтезе различных веществ, восприятии раздражений и в движении листьев.
Трихомы — различные по форме, строению и функциям выросты клеток эпидермы: волоски, чешуйки, щетинки и т.п. Их подразделяют на кроющие и железистые. Железистые трихомы, в отличие от кроющих, имеют клетки, выделяющие секрет. Кроющие волоски, образуя на растении шерстистый, войлочный или иной покров, отражают часть солнечных лучей и тем самым уменьшают транспирацию. Иногда волоски находятся только там, где расположены устьица, например, на нижней стороне листа мать-и-мачехи. У некоторых растений живые волоски увеличивают общую испаряющую поверхность, что способствует ускорению транспирации.
Размеры трихом значительно варьируют. Наиболее длинные трихомы (до 5-6 см) покрывают семена хлопчатника. Кроющие трихомы имеют форму простых одно или многоклеточных, разветвленных или звездчатых волосков. Кроющие трихомы могут длительное время оставаться живыми или быстро отмирать, заполняясь воздухом (рис. 2.10).
От трихом, возникающих только при участии эпидермальных клеток, отличаются эмергенцы, в образовании которых участвуют и более глубоко расположенные ткани субэпидермальных слоев (рис. 2.11).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом