
Известно, что фосфоролиз играет ключевую роль в мобилизации полисахаридов. (В тканях человека и животных советскими биохимиками Е. Л. Розенфельд и И. А. Поповой обнаружен также фермент γ-амилаза, катализирующий отщепление остатков глюкозы от молекулы гликогена по α-1,4-связи. Однако ведущая роль в расщеплении гликогена в клетках принадлежит фосфорилазам.) Фосфорилазы переводят полисахариды (в частности, гликоген) из запасной формы в метаболически активную форму; в присутствии фосфорилазы гликоген распадается с образованием фосфорного эфира глюкозы (глюкозо-1-фосфата) без предварительного расщепления на более крупные обломки молекулы полисахарида.
Реакция, катализируемая фосфорилазой, в общей форме выглядит так:
В этой реакции (C6H10O5)n означает полисахаридную цепь гликогена, a (C6H10O5)n-1 ту же, цепь, но укороченную на один глюкозный остаток.

На рис. 82 изображен ход распада гликогена до глюкозо-1-фосфата и участие в этом процессе цАМФ. Фермент фосфорилаза существует в двух формах, одна из которых (фосфорилаза «а») активна, в то время как другая (фосфорилаза «в») обычно неактивна. Обе формы могут диссоциировать на субъединицы. Фосфорилаза «в» состоит из двух субъединиц, а фосфорилаза «а» — из четырех. Превращение фосфорилазы «в» в фосфорилазу «а» осуществляется фосфорилированием белка по уравнению:
2 мол. фосфорилазы «в» + 4 АТФ ->
1 мол. фосфорилазы «а» + 4 АДФ
Катализируется эта реакция ферментом, который называется киназой фосфорилазы. Было найдено, что эта киназа может существовать как в активной, так и неактивной форме, причем неактивная киназа фосфорилазы превращается в активную под влиянием фермента протеинкиназы (киназа киназы фосфорилазы). Активная форма последней образуется при участии цАМФ. Как уже отмечалось , цАМФ в свою очередь образуется из АТФ под действием фермента аденилатциклазы. Эта реакция стимулируется, в частности, адреналином и глюкагоном. Увеличение содержания адреналина ведет по этой сложной цепи реакций к превращению фосфорилазы «в» в фосфорилазу «а» и, следовательно, к освобождению глюкозы в виде глюкозо-1-фосфата из запасного полисахарида гликогена. Обратное превращение фосфорилазы «а» в фосфорилазу «в» катализируется ферментом фосфатазой (эта реакция практически необратима).
Следует заметить, что фосфорилаза «а» отщепляет глюкозные остатки, начиная от периферического конца наружных ветвей молекулы гликогена, и при приближении к α (1—->6)-связям ее действие прекращается. Иными словами, фосфоролиз продолжается только до точек ветвления в молекуле гликогена. Фермент амило-1,6-глюкозидаза способен расщеплять (1—>6)-связь у точки ветвления, после чего фосфорилаза «а» вновь получает возможность действовать до тех пор, пока не дойдет до следующей точки ветвления, и т. д.
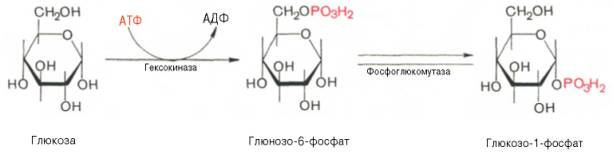
Образовавшийся в результате фосфоролиза глюкозо-1-фосфат превращается далее под действием фосфоглюкомутазы в глюкозо-6-фосфат:

Для того чтобы данная реакция протекала, необходима фосфорилированная форма фосфоглюкомутазы, т. е. ее активная форма, которая образуется в присутствии глюкозо-1,6-дифосфата. Таким образом, глюкозо-1,6-дифосфат в фосфоглюкомутазной реакции выполняет роль кофермента. (Глюкозо-1,6-дифосфат является продуктом следующей реакции: глюкозо-1-фосфат + АТФ глюкозо-1,6-дифосфат + АДФ).
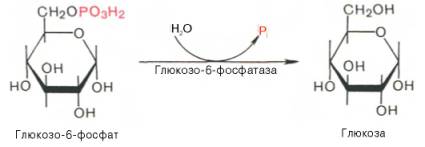
Образование свободной глюкозы из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы. (В отличие от печени в мышечной ткани глюкозо-6-фосфатазы нет).Данный фермент катализирует гидролитическое отщепление фосфата:

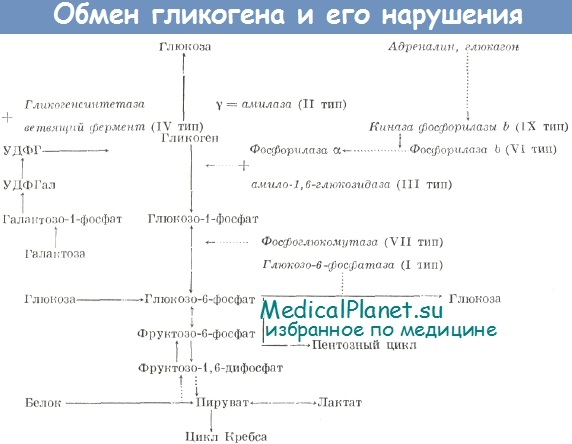
На рис. 83 изображены пути распада и синтеза гликогена.

Можно считать, что сохранение постоянства концентрации сахара в крови прежде всего есть результат одновременного протекания двух процессов: поступления глюкозы в кровь из печени и потребления ее из крови тканями, где она используется в первую очередь как энергетический материал.
В тканях (в том числе и в печени) существуют два основных пути распада глюкозы: анаэробный путь, который идет в отсутствие кислорода, и аэробный путь, для осуществления которого необходим кислород.
Распад гликогена (гликогенолиз)
Известно, что фосфоролитический распад играет ключевую роль в мобилизации полисахаридов.
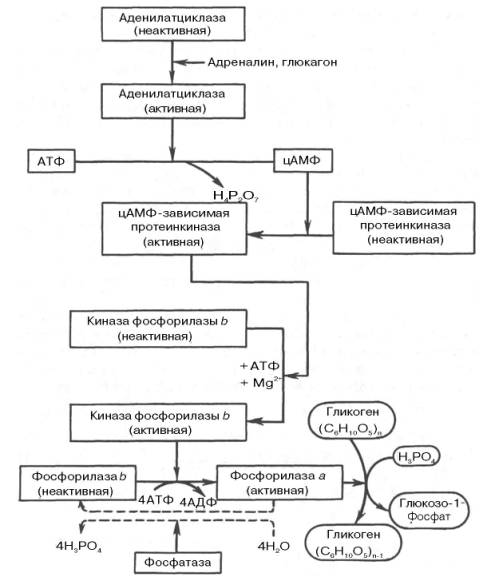
Рис. 10.1. Гормональная регуляция фосфоролитического отщепления остатка глюкозы от гликогена.
Читайте также: Стоун ткань для мебели
Фосфорилазы переводят полисахариды (в частности, гликоген) из запасной формы в метаболически активную форму; в присутствии фосфо-рилазы гликоген распадается с образованием фосфорного эфира глюкозы (глюкозо-1-фосфата) без предварительного расщепления на более крупные обломки молекулы полисахарида. В общей форме эту реакцию можно представить в следующем виде:
где (С6Н10О5)n означает полисахаридную цепь гликогена, а (С6Н10О5)n-1,– ту же цепь, но укороченную на один глюкозный остаток.
На рис. 10.1 изображены процесс распада гликогена до глюкозо-1-фосфата и участие в этом процессе цАМФ. Фермент фосфорилаза существует в двух формах, одна из которых (фосфорилаза а) активна, в то время как другая (фосфорилаза b) обычно неактивна. Обе формы могут диссоциировать на субъединицы. Фосфорилаза b состоит из двух субъединиц, а фосфорилаза а – из четырех. Превращение фосфо-рилазы b в фосфорилазу а осуществляется фосфорилированием белка:
2 Фосфорилаза b + 4 АТФ –> Фосфорилаза а + 4 АДФ.
Катализируется эта реакция ферментом, который называется киназой фосфорилазы b. Установлено, что эта киназа может существовать как в активной, так и в неактивной форме. Неактивная киназа фосфорилазы превращается в активную под влиянием фермента протеинкиназы (киназа киназы фосфорилазы), и не просто протеинкиназы, а цАМФ-зависимой протеинкиназы.
Активная форма последней образуется при участии цАМФ, которая в свою очередь образуется из АТФ под действием фермента аденилатцик-лазы, стимулируемой, в частности, адреналином и глюкагоном. Увеличение содержания адреналина в крови приводит в этой сложной цепи реакций к превращению фосфорилазы b в фосфорилазу а и, следовательно, к освобождению глюкозы в виде глюкозо-1-фосфата из запасного полисахарида гликогена. Обратное превращение фосфорилазы а в фосфорилазу b катализируется ферментом фосфатазой (эта реакция практически необратима).
Образовавшийся в результате фосфоролитического распада гликогена глюкозо-1-фосфат превращается под действием фосфоглюкомутазы в глюкозо-6-фосфат. Для осуществления данной реакции необходима фосфо-рилированная форма фосфоглюкомутазы, т.е. ее активная форма, которая образуется, как отмечалось, в присутствии глюкозо-1,6-бисфосфата .

Образование свободной глюкозы из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы. Данный фермент катализирует гидролитическое отщепление фосфата:
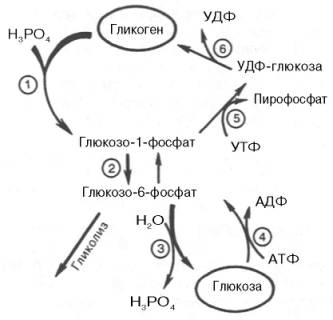
Жирными стрелками указан путь распада, тонкими — путь синтеза. Цифрами обозначены ферменты: 1 — фосфорилаза; 2 — фос-фоглюкомутаза; 3 — глюкозо-6-фосфатаза; 4 — гексокиназа (глюкокиназа); 5 — глюко-зо-1-фосфат-уридилтрансфераза; 6 — глико-генсинтаза.
Заметим, что фосфорилированная глюкоза в противоположность неэте-рифицированной глюкозе не может легко диффундировать из клеток. Печень содержит гидролитический фермент глюкозо-6-фосфатазу, который и обеспечивает возможность быстрого выхода глюкозы из этого органа. В мышечной ткани глюкозо-6-фосфатаза практически отсутствует.
На рис. 10.2 отражены представления о путях распада и синтеза гликогена в печени.
Можно считать, что сохранение постоянства концентрации глюкозы в крови является результатом одновременного протекания двух процессов: поступления глюкозы в кровь из печени и потребления ее из крови тканями, где она используется в первую очередь как энергетический материал.
В тканях (в том числе в печени) распад глюкозы происходит двумя основными путями: анаэробным (при отсутствии кислорода) и аэробным, для осуществления которого необходим кислород.
Синтез гликогена (гликогенез)
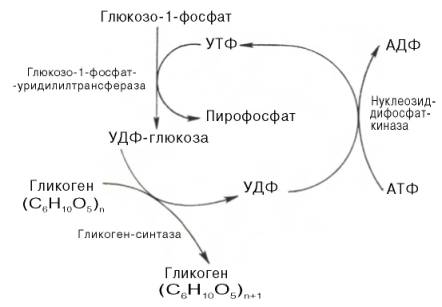
Прежде всего глюкоза подвергается фосфорилированию при участии фермента гексокиназы, а в печени – и глюкокиназы. Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фос-фат:
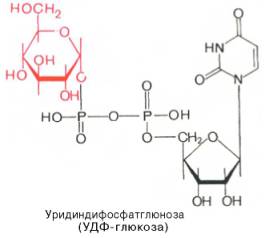
Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена. На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ (уридинтрифосфат), образуя уридиндифосфатглюкозу (УДФ-глюкоза) и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорилаза):
Глюкозо-1-фосфат + УТФ УДФ-глюкоза + Пирофосфат.
На второй стадии – стадии образования гликогена – происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную цепь гликогена («затравочное» количество). При этом образуется α-(1–>4)-связь между первым атомом углерода добавляемого остатка глюкозы и 4-гидроксильной группой остатка глюкозы цепи. Эта реакция катализируется ферментом гликогенсинтазой. Необходимо еще раз подчеркнуть, что реакция, катализируемая гликогенсинтазой, возможна только при условии, что полисахаридная цепь уже содержит более 4 остатков D-глю-козы.
Читайте также: Детский пенал из ткани
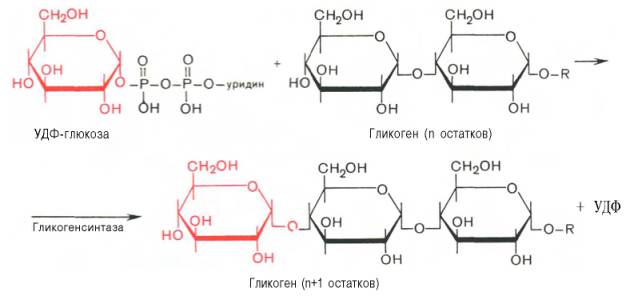
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и таким образом весь цикл превращений глюкозо-1-фосфата начинается сначала.
В целом образование α-1,4-глюкозидной ветви («амилозной» ветви) гликогена можно представить в виде следующей схемы:
Установлено, что гликогенсинтаза неспособна катализировать образование α-(1–>6)-связи, имеющейся в точках ветвления гликогена. Этот процесс катализирует специальный фермент, получивший название гли-когенветвящего фермента, или амило-(1–>4)–>(1–>6)-трансглюкозидазы. Последний катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6 или 7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксиль-ную группу остатка глюкозы той же или другой цепи гликогена. В результате образуется новая боковая цепь.
Ветвление повышает растворимость гликогена. Кроме того, благодаря ветвлению создается большое количество невосстанавливающих концевых остатков, которые являются местами действия гликогенфосфорилазы и гликогенсинтазы.
Таким образом, ветвление увеличивает скорость синтеза и расщепления гликогена.
Благодаря способности к отложению гликогена (главным образом в печени и мышцах и в меньшей степени в других органах и тканях) создаются условия для накопления в норме некоторого резерва углеводов. При повышении энерготрат в организме в результате возбуждения ЦНС обычно происходят усиление распада гликогена и образование глюкозы.
Помимо непосредственной передачи нервных импульсов к эффекторным органам и тканям, при возбуждении ЦНС повышаются функции ряда желез внутренней секреции (мозговое вещество надпочечников, щитовидная железа, гипофиз и др.), гормоны которых активируют распад гликогена, прежде всего в печени и мышцах (см. главу 8).
Как отмечалось, эффект катехоламинов в значительной мере опосредован действием цАМФ, который активирует протеинкиназы тканей. При участии последних происходит фосфорилирование ряда белков, в том числе гликогенсинтазы и фосфорилазы b – ферментов, участвующих в обмене углеводов. Фосфорилированный фермент гликогенсинтаза сам по себе малоактивен или полностью неактивен, но в значительной мере активируется положительным модулятором глюкозо-6-фосфатом, который увеличивает Vmaxфермента. Эта форма гликогенсинтазы называется D-формой, или зависимой (dependent) формой, поскольку ее активность зависит от глюкозо-6-фосфата. Дефосфорилированная форма гликоген-синтазы, называемая также I-формой, или независимой (independent) формой, активна и в отсутствие глюкозо-6-фосфата.
Таким образом, адреналин оказывает двойное действие на обмен углеводов: ингибирует синтез гликогена из УДФ-глюкозы, поскольку для проявления максимальной активности D-формы гликогенсинтазы нужны очень высокие концентрации глюкозо-6-фосфата, и ускоряет распад гликогена, так как способствует образованию активной фосфорилазы а. В целом суммарный результат действия адреналина состоит в ускорении превращения гликогена в глюкозу.
Распад гликогена в тканях до глюкозо 1 фосфат
а) Инсулин увеличивает облегченную диффузию глюкозы. Скорость транспорта глюкозы, как и транспорта других моносахаридов, существенно увеличивается инсулином. Если поджелудочная железа продуцирует большие количества инсулина, скорость транспорта глюкозы в большинстве клеток возрастает более чем в 10 раз по сравнению со скоростью транспорта глюкозы при отсутствии инсулина. Напротив, при отсутствии инсулина количество глюкозы, которое может диффундировать внутрь большинства клеток, за исключением клеток мозга и печени, столь мало, что не в состоянии обеспечить нормальный уровень энергетических потребностей.
Скорость потребления глюкозы большинством клеток находится под контролирующим влиянием скорости продукции инсулина поджелудочной железой. Функции инсулина и его регуляторные влияния на обмен углеводов подробно изложены в отдельной статье на сайте (просим вас пользоваться формой поиска выше).
б) Фосфорилирование глюкозы. Как только глюкоза попадает в клетки, она связывается с фосфатными радикалами в соответствии со следующей схемой реакции:
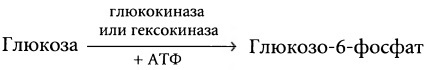
Фосфорилирование осуществляется главным образом ферментом глюкокиназой в печени или гексокиназой в большинстве других клеток. Фосфорилирование глюкозы является почти полностью необратимой реакцией, исключая клетки печени, эпителиоциты почечного тубулярного аппарата и клетки кишечного эпителия, в которых присутствует другой фермент — глюкофосфорилаза. Будучи активирована, она может сделать реакцию обратимой. В большинстве тканей организма фосфорилирование служит способом захвата глюкозы клетками. Это происходит в связи со способностью глюкозы немедленно связываться с фосфатом, а в такой форме она не может выходить обратно из клетки, кроме некоторых особых случаев, в частности из клеток печени, которые располагают ферментом фосфатазой.
Читайте также: Вышивка лентами сумок из ткани
в) Гликоген запасается в печени и мышцах. После поступления внутрь клетки глюкоза практически немедленно используется клеткой для энергетических целей либо запасается в виде гликогена, который является крупным полимером глюкозы.
Все клетки организма способны запасать какие-то количества гликогена, но особенно большие его количества депонируются клетками печени, которая может запасать гликоген в количествах, составляющих от 5 до 8% веса этого органа, или мышечными клетками, содержание гликогена в которых составляет от 1 до 3%. Молекула гликогена может полимеризоваться так, что в состоянии иметь практически любую молекулярную массу; в среднем молекулярная масса гликогена составляет около 5 млн. В большинстве случаев гликоген, осаждаясь, образует крупные гранулы.
Превращение моносахаридов в преципитирующее соединение с большой молекулярной массой (гликоген) дает возможность запасать большие количества углеводов без заметного изменения осмотического давления во внутриклеточном пространстве. Высокая концентрация растворимых моносахаридов с низкой молекулярной массой могла бы приводить к катастрофическим последствиям для клеток в связи с формированием огромного градиента осмотического давления по обе стороны клеточной мембраны.
г) Гликогенез — процесс образования гликогена. Химические реакции образования гликогена показаны на рисунке ниже.
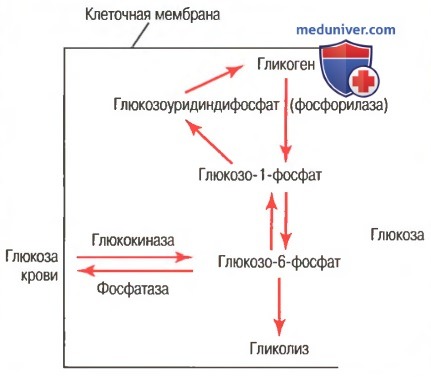
Химические реакции глюкогенеза и гликогенолиза. Показаны также взаимные превращения глюкозы крови и гликогена печени (фосфатаза требуется для высвобождения глюкозы из клеток и представлена в клетках печени, но отсутствует в большинстве других клеток)
На рисунке видно, что глюкозо-6-фосфат становится глюкозо-1-фосфатом, который затем превращается в глюкозоуридинфосфат, в итоге образующий гликоген. Для этих превращений необходимы специфические ферменты. Кроме того, и другие моносахариды, превращаясь в глюкозу, могут участвовать в образовании гликогена. Более мелкие соединения, включая молочную кислоту, глицерол, пировиноградную кислоту и некоторые дезаминированные аминокислоты, также могут превращаться в глюкозу или близкие ей соединения и затем становиться гликогеном.
д) Гликогенолиз — извлечение гликогена из депо. Процесс расщепления гликогена, хранящегося в клетках, который сопровождается высвобождением глюкозы, называют гликогенолизом. Затем глюкоза может использоваться в целях получения энергии. Гликогенолиз невозможен без реакций, обратных реакциям получения гликогена, при этом каждая вновь отщепляющаяся от гликогена молекула глюкозы подвергается фосфорилированию, катализируемому ферментом фосфорилазой. В состоянии покоя фосфорилаза пребывает в неактивном состоянии, поэтому гликоген может храниться в депо. Когда появляется необходимость получить глюкозу из гликогена, прежде всего должна активироваться фосфорилаза. Это может достигаться несколькими путями.
е) Активация фосфорилазы адреналином или глюкагоном. Два гормона — адреналин и глюкагон — могут активировать фосфорилазу и таким образом ускорять процессы гликогенолиза. Начальные моменты влияний этих гормонов связаны с образованием в клетках циклического аденозинмонофосфатау который затем запускает каскад химических реакций, активирующих фосфорилазу.
Адреналин выделяется из мозгового вещества надпочечников под влиянием активации симпатической нервной системы, поэтому одна из ее функций заключается в обеспечении обменных процессов. Эффект адреналина особенно заметен в отношении клеток печени и скелетных мышц, что обеспечивает наряду с влияниями симпатической нервной системы готовность организма к действию.
Глюкагон — гормон, выделяемый альфа-клетками поджелудочной железы, когда концентрация глюкозы в крови снижается до слишком низких значений. Он стимулирует образование циклического АМФ главным образом в клетках печени, что, в свою очередь, обеспечивает превращение в печени гликогена в глюкозу и ее высвобождение в кровь, повышая таким образом концентрацию глюкозы в крови.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом