

В любом случае в первую очередь происходит активация глицерина.
Она похожа на активацию углеводов.
Распад глицерина по пути к углеводам
Если глицерин распадается по пути к углеводам, то происходит дегидрирование.
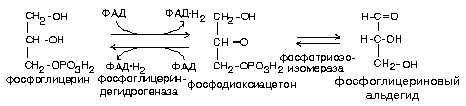
Митохондриальная фосфоглицериндегидрогеназа содержит в качестве небелковой части ФАД, а цитоплазматическая — НАД. В митохондриях отщепляемый водород переносится по укороченной цепи митохондриального окисления, и образуется 2 молекулы АТФ (фосфоглицерин (ФГА) — субстрат укороченной цепи).
Для фосфоглицеринового альдегида существует два варианта дальнейших превращений:
ФГА может окисляться в ГБФ-пути до СО2 и Н2О с образованием 21 молекулы АТФ.
ФГА может вступить в реакции гликонеогенеза с образованием углеводов — глюкозы или гликогена (смотрите тему «Метаболизм углеводов и его регуляция»).
Пути использования жирных кислот.
Жирные кислоты могут вступать в реакции только после активации. Активация жирных кислот принципиально отличается от активации углеводов.
Реакция начинается с переноса от АТФ не фосфата, а АМФ, с образованием промежуточного продукта — ациладенилата. Затем с участием HS-KoA отщепляется АМФ, и образуется активная форма любой жирной кислоты – АЦИЛ-КоА.
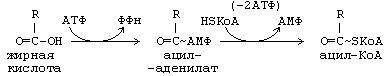
Образовавшийся АМФ не может превратиться в АТФ. Поэтому протекает еще одна реакция, и тоже – с затратой АТФ:
Как видно, распад 1 АТФ до АМФ энергетически равен распаду 2-х АТФ до 2-х АДФ. Поэтому затраты энергии на активацию жирной кислоты составляют 2 АТФ на одну молекулу жирной кислоты.
Для активной жирной кислоты, как и для глицерина, возможны два пути метаболических превращений:
1. Синтез жира или других липидов.
2. Катаболизм до Ацетил-КоА. Этот процесс называют БЕТА-ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ.
Катаболизм жирных кислот
Активация ЖК происходит в цитоплазме, а бета-окисление — В МИТОХОНДРИЯХ.
Ацил-КоА не может проходить через мембрану митохондрий. Поэтому имеется специальный механизм транспорта ЖК из цитоплазмы в митохондрию при участии вещества «КАРНИТИН». Во внутренней мембране митохондрий есть специальный транспортный белок, обеспечивающий перенос. Благодаря этому ацилкарнитин легко проникает через мембрану митохондрий.
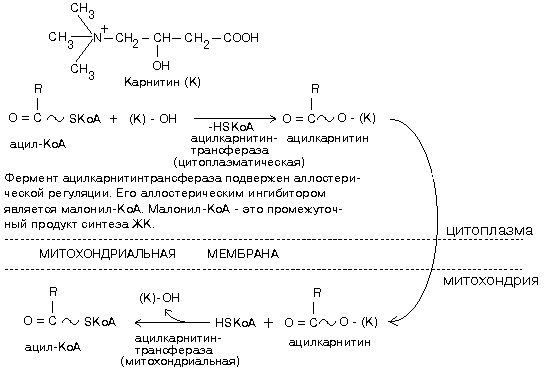
По строению цитоплазматическая и митохондриальная карнитинацилтрасферазы различны, отличаются они друг от друга и кинетическими характеристиками. Vmaxцитоплазматической ацилкарнитинтрансферазы ниже, чемVmaxмитохондриального фермента, а также нижеVmaxферментов-окисления. Поэтому цитоплазматическая ацилкарнитинтрансфераза является ключевым ферментом распада жирных кислот.
Если жирная кислота попадает в митохондрию, то она обязательно подвергнется катаболизму до ацетил-КоА.
Процесс -окисления является циклическим. За каждый оборот цикла от жирной кислоты отщепляется 2 углеродных атома в виде ацетильного остатка.
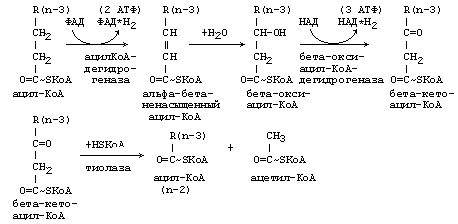
После этого укороченный на 2 углеродных атома ацил-КоА снова подвергается окислению (вступает в новый цикл реакций -окисления). Образующийся Ацетил-КоА может дальше вступить в цикл трикарбоновых кислот.
Нужно уметь рассчитывать энергетический выход при распаде жирных кислот. Представленная формула верна для любой насыщенной жирной кислоты, содержащей n углеродных атомов:
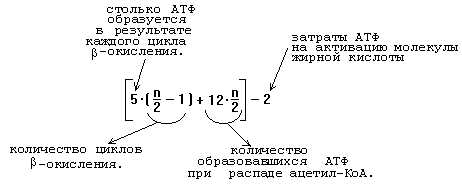
При распаде ненасыщенных жирных кислот образуется меньше АТФ. Каждая двойная связь в жирной кислоте — это потеря 2-х молекул АТФ.
-окисление наиболее интенсивно протекает в мышечной ткани, почках, печени.
В результате -окисления ЖК образуется Ацетил-КоА. Скорость окисления определяется скоростью процессов липолиза. Ускорение липолиза характерно для состояния углеводного голодания и интенсивной мышечной работы. Ускорение -окисления наблюдается во многих тканях, в том числе и в печени. В печени образуется больше Ацетил-КоА, чем ей требуется. Печень — «орган-альтруист» и поэтому печень отправляет глюкозу в другие ткани. Печень стремится направить в другие ткани и свой собственный Ацетил-КоА, но не может, так как для Ацетил-КоА клеточные мембраны непроницаемы. Поэтому в печени из Ацетил-КоА синтезируются специальные вещества, которые называются «КЕТОНОВЫЕ ТЕЛА».
КЕТОНОВЫЕ ТЕЛА — ЭТО ОСОБАЯ ТРАНСПОРТНАЯ ФОРМА АЦЕТИЛ-КоА!
66. Окисление глицерина в тканях. Энергетический эффект этого процесса.
В результате гидролиза жира образуются общие метаболиты: глицерины и ВЖК, окисление которых сопровождается образованием конечных продуктов -воды и углекислого газа -и выделением энергии в форме АТФ. Окисление глицеринов в тканях тесно связано с ГЛИКОЛИЗОМ, в который вовлекаются метаболиты обмена глицерина по следующей схеме:
Читайте также: Геотекстиль что это за ткань

Т.о. при окислении глицерина образовались конечные продукты:
3. Альфа-КЕТОГЛУТАРАТА АТФ на этапе превращения:
3. 1,3-ДИФОСФОГЛИЦЕРИНОВОЙ К-ТЫ (СУБСТРАТНОЕФОСФОРИЛИРОВАНИЕ)
4. 2-ФОСФОЕНОЛПИРУВАТА (СУБСТРАТНОЕ ФОСФОРИЛИРОВАНИЕ)
8. СУКЦИНИЛ-КОА (СУБСТРАТНОЕ ФОСФОРИЛИРОВАНИЕ)
67. Биосинтез высших жирных кислот в тканях. Биосинтез жиров в печени и жировой ткани.
Биосинтез ВЖК протекает в ЭПС клеток. Исходным материалом для синтеза заменимых ВЖК является АЦЁТИЛ-КОА.
Условия для биосинтеза ВЖК:
1.Наличие АТФ, СО2, Н2О, НАДФ*Н2, поступающий из ГЕКСОЗОМОНОФОСФАТНОГО пути превращения глюкозы.
2.Наличие специальных белков-переносчиков (HS -АПБ).
3.Наличие специальных ферментов синтеза.
Процесс биосинтеза циклический. Каждый цикл включает в себя 6 этапов. АЦЕТИЛ-КОА используется на первом этапе, как «затравка» синтеза.
Первый этап. Этап образования 3-углеродного соединения — МАЛОНИЛ-КОА.
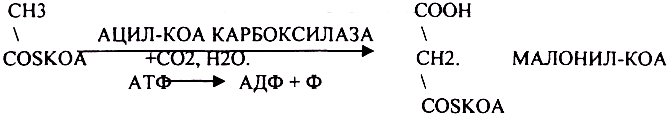
Второй этап. Этап переноса МАЛОНИЛА и АЦЕТИЛА на специальные белки — HS-АПБ.

Третий этап. Этап конденсации МАЛОНИЛА-АПБ и АЦЕТИЛА-АПБ с участием ТРАНСФЕР АЗЫ — СИНТАЗЫ.
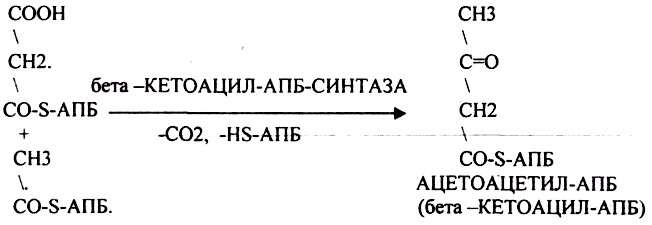
Четвёртый этап. Этап восстановления бета -КЕТОАЦИЛ-АПБ.

Пятый этап. Этап ДЕГИДРАТАЦИИ бета-ГИДРОКСИАЦИЛ-АПБ.

Шестой этап. Этап восстановления ЕНОИЛАЦИЛ-АПБ.
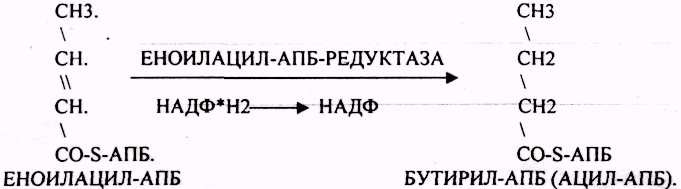 Т.о. завершается 1 цикл синтеза ВЖК образованием масляной кислоты (БУТИРИЛ-АПБ). В дальнейшем последовательно и циклично к 4-углеродному фрагменту будут присоединяться молекулы МАЛОНИЛ-КОА. Для подсчёта количества циклов синтеза ВЖК, подсчёта молекул АТФ, МАЛОНИЛ-КОА можно пользоваться формулой: N/2 — 1, где N-это число углеродных звеньев в молекуле ВЖК. Завершается биосинтез любой ВЖК в тканях ДЕАЦИЛАЗНОЙ реакцией.
Т.о. завершается 1 цикл синтеза ВЖК образованием масляной кислоты (БУТИРИЛ-АПБ). В дальнейшем последовательно и циклично к 4-углеродному фрагменту будут присоединяться молекулы МАЛОНИЛ-КОА. Для подсчёта количества циклов синтеза ВЖК, подсчёта молекул АТФ, МАЛОНИЛ-КОА можно пользоваться формулой: N/2 — 1, где N-это число углеродных звеньев в молекуле ВЖК. Завершается биосинтез любой ВЖК в тканях ДЕАЦИЛАЗНОЙ реакцией.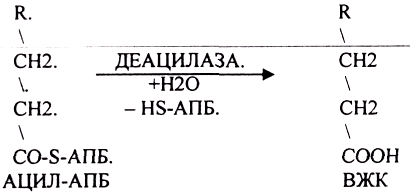
Так в организме человека происходит синтез всех заменимых ВЖК — всех предельных и непредельных ВЖК, имеющих одну двойную связь.
51. Холестерол. Его химическое строение, биосинтез и биологическая роль. Причины гиперхолестеринемии.
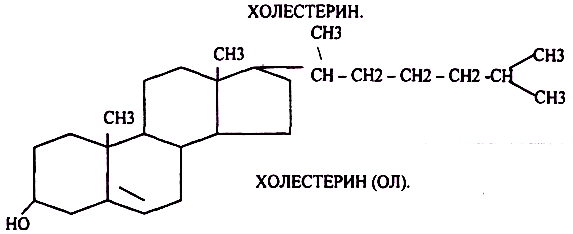
Холестерин является одноатомным циклическим спиртом, который в тканях легко образует ХОЛЕСТЕРИДЫ. В организм человека поступает в составе пищи и синтезируется г.о. в печени, тонком отделе кишечника и коже.
Биологическая роль холестерина:
1.Структурная. Свободный холестерин является, обязательным структурным компонентом мембран клеток.
2.Метаболическая. Холестерин является предшественником биологически активных веществ: витамина D3,СТЕРОИДНЫХ гормонов (АНДРОГЕНОВ, ЭСТРОГЕНОВ, КОРТИКОИДОВ) При окислении холестерина в печени при участии ЦИТОХРОМА Р-450 образуются желчные кислоты. В свободном виде холестерин транспортируется по организму с помощью транспортных ЛИПОПРОТЕИНОВ крови. Источники холестерина:
1. Пища. За сутки в организм взрослого человека поступает 0,3гр. холестерина.
2. У человека в среднем с массой 65-70кг за сутки синтезируется 3.5 -4,2гр. холестерина. Печень занимает главное место в синтезе холестерина (85%), холестерин синтезируется в кишечнике (10%) и коже (5%). Процесс биосинтеза многоступенчат:

Образовавшийся в результате распада мембранных ЛИПИДОВ, а также излишки холестерина с помощью ЛПВП доставляется для окисления в печень, и в составе желчных кислот удаляется из организма с каловыми массами. При поражении печени и кишечника нарушается образование и транспорт ЛП крови. При поражении печени и желчевыводяицих путей нарушается образование и экскреция желчных кислот, участвующих в переваривании жиров пищи. В случае нарушения оттока желчи происходит насыщение её холестерином, что ведёт в этих условиях к застою и образованию холестериновых камней. Развивается ЖЕЛЧЕКАМЕННАЯ БОЛЕЗНЬ. В крови отмечается ГИПЕРХОЛЕСТЕРИНЕМИЯ.
52. Основные проявления патологии липидного обмена и возможные причины их возникновения на различных этапах обмена веществ. Образование кетоновых тел в тканях. Кетоацидоз. Биологическое значение кетоновых тел.
1 .На этапе поступления жиров с пищей:
A. Обильная жирная пища на фоне ГИПОДИНАМИИ приводит к развитию АЛИМЕНТАРНОГО ОЖИРЕНИЯ.
B. Недостаточное поступление жиров или их отсутствие приводит к ГИПО- и АВИТАМИНОЗАМ A, D, Е, К. Могут развиваться ДЕРМАТИТЫ, СКЛЕРОЗ сосудов. Также нарушается процесс синтеза ПРОСТАГЛАНДИНОВ.
Читайте также: Техника выполнения поделки из ткани
C. Недостаточное поступление с пищей ЛИПОТРОПНЫХ(холин, серин, инозит, витамины В12, В6) веществ приводит к развитию жировой инфильтрации тканей.
A. При поражении печени и кишечника нарушается образование и транспорт ЛП крови.
B. При поражении печени и желчевыводяицих путей нарушается образование и экскреция желчных кислот, участвующих в переваривании жиров пищи. Развивается ЖКБ. В крови отмечается ГИПЕРХОЛЕСТЕРИНЕМИЯ.
C. Если поражается слизистая оболочка кишечника и нарушается выработка и поступление ферментов ПЖЖ, содержание жира в кале увеличивается. Если содержание жира будет превышать 50%, развивается СТЕАТОРЕЯ. Кал становится бесцветным.
D. Наиболее часто в последнее время среди населения встречается поражение бета -клеток ПЖЖ, что ведет к развитию сахарного диабета, который сопровождается интенсивным окислением в клетках белков и жиров. В крови у таких больных отмечается ГИПЕРКЕТОНЕМИЯ, ГИПЕРХОЛЕСТЕРИНЕМИЯ. Кетоновые тела и холестерин синтезируются из АЦЕТИЛ-КОА.
3.На этапе обмена холестерина наиболее частым заболеванием является АТЕРОСКЛЕРОЗ. Болезнь развивается тогда, когда между клетками тканей и ЛП крови растёт содержание АТЕРОГЕННЫХ ФРАКЦИЙ и понижается содержание ЛПВП, назначение которых удалять холестерин из клеток тканей в печень для последующего его окисления. Все ЛП за исключением ХИЛОМИКРОНОВ быстро метаболизируются. ЛПНП задерживаются в сосудистой стенке. Они содержат много ТРИГЛИЦЕРИДОВ и ХОЛЕСТЕРИНА. Они, фагоцитируясь, разрушаются ферментами ЛИЗОСОМ, за исключением холестерина. Он накапливается в клетке в большом количестве. Холестерин откладывается в межклеточном пространстве и инкапсулируется соединительной тканью. В сосудах образуются АТЕРОСКЛЕРОТИЧЕСКИЕ БЛЯШКИ.

Кетоновые тела (не более 0,1 г/л) — ацетон, ацетоуксусная кислота, бета -гидроксимасляная кислота. При дефиците углеводов в клетке жиры не могут полностью окисляться, и избыток ацетил-КоА компенсируется образованием кетоновых тел. Опасно в отношении КЕТОАЦИДОЗА.
11.2. Метаболизм глицерина
Обмен глицерина тесно связан с гликолизом, в который вовлекаются метаболиты глицерина по следующей схеме:
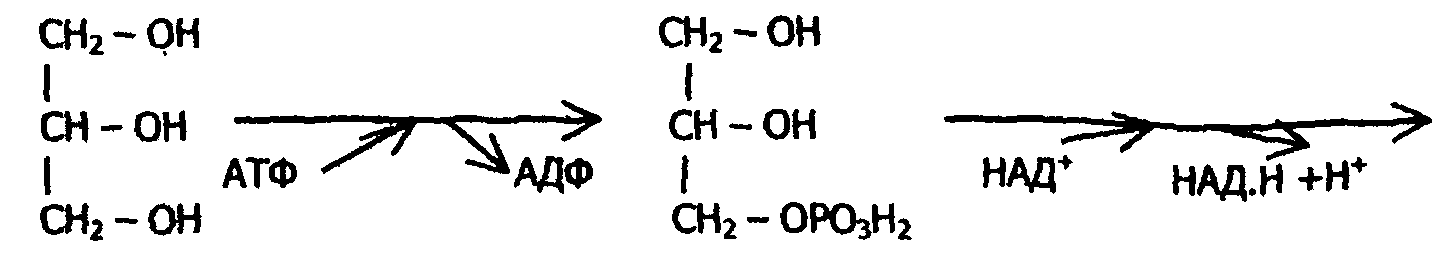

Превращение одной молекулы глицерина дает одну молекулу АТФ в анаэробных условиях и 19 молекул АТФ в аэробных. Глицерин как энергетический материал используется практически всеми органами и тканями.
11.3. Метаболизм жирных кислот
В триацилглицеринах (жирах) жировой ткани человека в основном содержатся следующие жирные кислоты: миристиновая (3%), пальмитиновая (20%), стеариновая (5%), пальмитоолеиновая (5%), олеиновая (55%), линолевая (10%), арахидоновая (0,2%). В значительных количествах эти жирные кислоты содержатся и в других липидах, но жирнокислотный состав гликолипидов и фосфолипидов клеточных мембран гораздо более разнообразен. Особенно много характерных жирных кислот найдено в сложных липидах нервных клеток.
Источниками жирных кислот организма служат липиды пищи (главным образом жиры) и синтез жирных кислот из углеводов.
Расходуются жирные кислоты в основном по трем направлениям (рис.33):
— включаются в состав резервных жиров;
— включаются в состав структурных липидов;
— окисляются до углекислого газа и воды с использованием выделяющейся при этом энергии для синтеза АТФ.

Рис. 33. Метаболизм жирных кислот
Все превращения сложных жирных кислот в клетках начинаются с образования Ацил-КоА (активация жирных кислот):
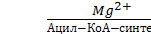
СН3-(СН2)n-СН2-СН2-СООН + HSKoA + АТФ

СН3-(СН2)n-СН2-СН2-С
Содержащаяся в Ацил-КоА связь C
S является макроэргической, поэтому данный процесс и рассматривают как активацию кислоты.
Дальнейший катаболизм жирных кислот можно разделить на три стадии:
1) β-окисление — специфический для жирных кислот путь метаболизма, завершающийся превращением молекулы жирной кислоты в несколько молекул Ацетил-КоА;
2) цикл Кребса, в котором окисляются ацетильные остатки;
3) Митохондриальная дыхательная цепь.
Процесс активации жирных кислот протекает в цитоплазме, а β-окисление активированных кислот происходит в матриксе митохондрий при участии мультиферментного комплекса. Мембрана митохондрий непроницаема для жирных кислот; их перенос происходит при участии карнитина:
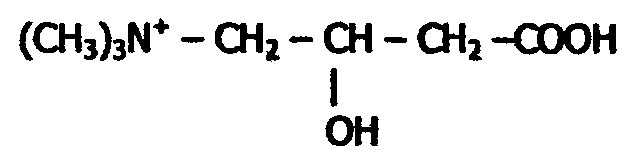
При действии карнитин-ацилтрансферазы к спиртовой группе карнитина присоединяется ацильный остаток жирной кислоты (сложно-эфирной связью):

Образующийся ацилкарнитин может диффундировать в митохондрию, где происходит обратная реакция с образованием Ацил-КоА.
Читайте также: Для изготовления пальто подойдут ткани с чем
В матриксе митохондрий происходит β-окисление поступившего Ацил-КоА. При β-окислении окисляется группа –СН2 — в β-положении по отношению к группе -СО-:


Новый Ацил-КоА вновь подвергается β-окислению. Многократное повторение этого процесса приводит к полному распаду жирной кислоты до Ацетил-КоА. Напиример, молекула пальмитиновой кислоты, содержащая 16 атомов углерода, превращаясь в 8 молекул Ацетил-КоА за 7 циклов β-окисления:

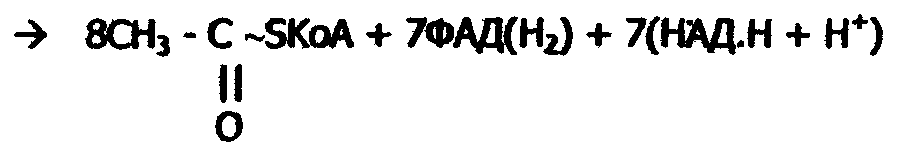
Окисление кислот с нечетным числом атомов углерода и ненасыщенных кислот имеет свои особенности.
В случае кислот с нечетным количеством атомов углерода наряду с обычными продуктами окисления образуется одна молекула пропионил-КоА (CH3-CH2-CO
SKoA) на молекулу окисленной жирной кислоты. Пропионил-КоА окисляется по особому пути:

Образующийся сукцинил-КоА поступает в цикл Кребса.
Особенности окисления ненасыщенных жирных кислот определяются положением и числом двойных связей в их молекулах. Окисление идет обычным путем, если каждая двойная связь имеет трансконфигурацию. В противном случае в реакциях участвует дополнительный фермент, изменяющий конфигурацию групп атомов относительно двойной связи из цис- в транс-, далее окисление идет так же, как у насыщенных кислот. Следует отметить, что скорость окисления ненасыщенных жирных кислот выше, чем насыщенных. Например, по сравнению с окислением стеариновой кислоты скорость окисления олеиновой выше в 11 раз, линолевой — в 114, линоленовой — в 170 раз, а арахидоновой — почти в 200 раз.
Энергетическая ценность жирной кислоты с четным числом атомов углерода рассчитывается следующим образом. Если жирная кислота содержит 2n атомов углерода, то при полном ее окислении образуется n молекул ацетил-КоА и по ( n-1 ) молекул ФАД(Н2) и (НАД.Н + Н + ). Окисление ФАД(Н2) дает 2 АТФ, а (НАД.Н+Н + )-3 АТФ, то есть вместе — 5 АТФ или, в общем виде, 5(n-1) АТФ. Полное сгорание одной молекулы ацетил-КоА дает 12 АТФ, значит n молекул обеспечивают образование 12n АТФ. Учитывая, что 1 АТФ тратится на активирование кислоты, полный баланс АТФ при окислении жирной кислоты с четным числом атомов углерода можно выразить формулой:
где n=m/2 (m- число атомов углерода в кислоте).
Например, полный выход АТФ при окислении одной молекулы пальмитиновой кислоты составляет 130 молекул.
Энергетическая ценность жирных кислот выше, чем, например, глюкозы. Так, полное окисление капроновой кислоты, имеющей то же число атомов углерода, что и глюкоза, дает 45 молекул АТФ (глюкоза дает 38 молекул АТФ). Однако для сгорания в цикле Кребса образующихся при β-окислении молекул ацетил-КоА требуется достаточное количество оксалоацетата. В этом отношении углеводы имеют преимущество перед жирными кислотами, так как при их распаде образуется пируват, являющийся источником образования не только ацетил-КоА, но и оксалоацетата, то есть облегчается превращение ацетил-КоА в цикле Кребса. Не случайно в биохимической литературе бытовало выражение: «жиры сгорают в пламени углеводов», поскольку образующийся уже в гликолизе АТФ может использоваться для активирования жирных кислот в цитоплазме, а образующийся из пирувата оксалоацетат обеспечивает включение ацетил-КоА в цикл Кребса.
β-Окисление жирных кислот происходит во многих тканях, но особенно значительна роль этого источника энергии в скелетных мышцах при большой физической нагрузке, а также в сердечной мышце и в почках. Сердечная мышца около 70% поглощаемого кислорода использует для окисления жирных кислот, а нервная ткань, например, вообще не использует этот источник энергии.
Часть Ацетил-КоА минует цикл Кребса и расходуется на синтез стероидов, прежде всего холестерина, и жирных кислот в цитоплазме клеток различных органов и тканей. Холестерин в наибольшей степени синтезируется в печени (80%), а также в стенках тонкого кишечника (10%)и в клетках кожи (5%). За сутки образуется 1 г холестерина в организме, тогда как с пищей в организм поступает 0,1-0,3 г холестерина, всего 8 тканях организма холестерина приблизительно 140 г, на втором месте группа стероидов желчных кислот — приблизительно 5 г.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом