
1. Переваривание липидов и всасывание продуктов их распада в ЖКТ.
2. Распад липидов в тканях: а) превращение глицерина в тканях;
б) превращение жирных кислот в тканях.
3. Биосинтез триглицеридов в организме.
В ротовой полости жиры не перевариваются, так как нет условий (ферментов) для переваривания. В желудке идёт частичный гидролиз жиров. Основное переваривание идёт в тонком кишечнике. При участии фермента липазы, вырабатываемой поджелудочной железой, и секрета печени – желчи. Сначала отделяются ЖК по концам молекулы жира из α – положения, далее в β.
| липаза |
| липаза |
Липаза активизируется желчью, в состав которой входят желчные кислоты: холевая, дезоксихолевая, гликохолевая, литохолевая – по строению они близки к холестерину. Холевые кислоты придают горький вкус и грязный цвет желчи.
Роль желчи в переваривании жира:
1. Желчь – активатор липазы.
3. Желчь способствует всасыванию жирных кислот, образующихся при гидролизе жира.
Образующийся глицерин и жирные кислоты из кишечника поступают в кровь, доставляются в ткани, где претерпевают дальнейшее превращение.
В тканях есть тканевые липазы – липазы печени, легких, мышц. Активизация тканевых липаз идет засчет энергии АТФ. В тканях идет распад жира также на глицерин и жирные кислоты, которые подвергаются дальнейшему превращению.
а) Распад глицерина в тканях
Глицерин, независимо от того пойдет он на ресинтез жира или на дальнейший распад сначала активизируется или фосфорилируется за счет АТФ в присутствии киназы и затем через ряд промежуточных продуктов переходит в ПВК, аналогично гликолизу.
| киназа -АДФ |
| → | 1,3-диФГК | → | 3-ФГК | → | 2-ФГК | → | фосфо-ен | → | ПВК | → | ен-ПВК | → |
| COOH | ||||
| | | молочная кислота | |||
| C = O | ||||
| | | ||||
| CH3 | ацетил-КоА | → | цикл Кребса | (CO2+H2O) |
| ПВК |
Расчет энергетического эффекта распада глицерина в тканях:
ацетил- КоА → ц. Кребса 12 АТФ
Итого глицерин при окислении в тканях дает 19 АТФ.
б) Распад жирных кислот в тканях
В тканях при распаде жиров наряду с глицерином образуются жирные кислоты, которые подвержены дальнейшему превращению. Первые гипотезы в отношении механизма их распада были высказаны в 1904г Кнопом. Высшие ЖК предельные с четным числом атомов углерода распадаются в тканях преимуществом путем β-окисления. Это многостадийный процесс, который осуществляется специальными ферментами. Кноп обратил внимание на то, что в состав жиров входят жирные кислоты с четным числом атомов углерода. Окисление идет по углероду в β-положении. ЖК постепенно укорачиваются на двух углеродные фрагменты, в результате чего высшая кислота переходит в низкомолекулярную.
стеариновая к-та пальмитиновая миристиновая лауриновая
каприновая каприловая капроновая масляная уксусная
На первом этапе ЖК активизируется за счет соединения НS-КоА и энергии АТФ, в результате образуется активная форма жирной кислоты – ацил-КоА. Активная форма ЖК далее подвергается β- окислению. Например:
β-окисление капроновой кислоты
Таким образом, жирные кислоты в тканях окисляются до ацетил-КоА, который сгорает в цикле Кребса до CO2 и H2O.
капроновая кислота→3 ацетил-КоА 5-1 АТФ= 4+5=9 АТФ – за 2 оборота
3-ацетил-КоА → СО2 +Н2О 12АТФ*3= 36АТФ
При полном окислении капроновой кислоты образуется 45АТФ.
На синтез жира идут активная форма глицерина (глицерофосфат) и активная форма жирных кислот. Биосинтез жиров – это многостадийный процесс при участии специфических ферментов.
Жиры в своей основе содержат глицерин и набор жирных кислот. Если в состав жира входит одна и та же жирная кислота, то такой жир называется простым, а если набор жирных кислот разный, то такой жир называется смешанным. Например, простые жиры – тристеарин, триолеин, трипальметин; смешанный жир – пальмито-олео-стеарин.
Распад липидов в тканях
После всасывания в эпителий кишки свободные жирные кислоты и 2-моноглицериды вновь образуют триглицериды и вместе с фосфолипидами и холестеролом включаются в хиломикроны. Хиломикроны транспортируются с током лимфы через грудной проток в верхнюю полую вену, попадая таким образом в общий кровоток.
Внутри хиломикрона триглицериды гидролизуются липопротеинлипазой, что приводит к высвобождению жирных кислот на поверхности кровеносных капилляров в тканях. Это обусловливает транспорт жирных кислот в ткани и последующее образование остатков хиломикронов, обедненных триглицеридами. Эти остатки затем забирают эфиры холестерола из липопротеинов высокой плотности, и частицы быстро захватываются печенью. Данную систему транспорта жирных кислот пищевого происхождения называют экзогенной транспортной системой.
Также существует эндогенная транспортная система, предназначенная для внутриорганного транспорта жирных кислот, образованных в самом организме. Липиды транспортируются из печени в периферические ткани и обратно, а также переносятся из жировых депо к различным органам. Транспорт липидов от печени к периферическим тканям включает согласованные действия ЛПОНП, липопротеинов промежуточной плотности (ЛППП), липопротеинов низкой плотности (ЛПНП) и липопротеинов высокой плотности (ЛПВП). Частицы ЛПОНП подобно хиломикронам состоят из большого гидрофобного ядра, образованного триглицеридами и эфирами холестерола, и поверхностного липидного слоя, состоящего в основном из фосфолипидов и холестерола.
ЛПОНП синтезируются в печени, и отложение жира в периферических тканях является их основной функцией. После попадания в кровоток ЛПОНП подвергаются воздействию липопротеинлипазы, которая гидролизует триглицериды до свободных жирных кислот. Свободные жирные кислоты, происходящие из хиломикронов или ЛПОНП, могут быть использованы в качестве источников энергии, структурных компонентов фосфолипидных мембран или превращаться обратно в триглицериды и в таком виде сохраняться. Триглицериды хиломикронов и ЛПОНП также подвергаются гидролизу липазы печени.
Частицы ЛПОНП посредством гидролиза триглицеридов превращаются в более плотные, меньшие по размеру холестерол- и триглицерид-обогащенные остатки (ЛППП), которые удаляются из плазмы с помощью рецепторов печеночных липопротеинов или могут быть превращены в ЛПНП. ЛПНП являются основными липопротеинами-переносчиками холестерола.
Возврат липидов из периферических тканей в печень часто называют обратным транспортом холестерола. Частицы ЛПВП участвуют в этом процессе, забирая холестерол из тканей и других липопротеинов и перенося его в печень для последующей экскреции. Еще один вид транспортировки, существующей между органами, — перенос жирных кислот из жировых депо к органам для окисления.
Жирные кислоты, получаемые в основном в результате гидролиза триглицеридов жировой ткани, секретируются в плазму, где соединяются с альбумином. Связанные с альбумином жирные кислоты переносятся по градиенту концентрации в ткани с активным метаболизмом, где и используются преимущественно в качестве источников энергии.
В течение последних 20 лет только несколько исследований были посвящены вопросу транспорта липидов в перинатальном периоде (результаты этих исследований в данном издании не представлены). Очевидна необходимость более детального изучения этой проблемы.
Жирные кислоты используются в качестве строительного материала в составе липидов клеточной стенки, в качестве источников энергии, а также откладываются «про запас» в виде триглицеридов преимущественно в жировой ткани. Некоторые омега-6 и омега-3 ДЦПНЖК являются предшественниками биологически активных метаболитов, используемых в сигнальной системе клеток, регуляции генов и других метаболически активных системах.
Вопрос о роли ДЦПНЖК ARA и DHA в процессе роста и развития ребенка является одним из важнейших вопросов в исследованиях, проводимых в области детской нутрициологии в течение последних двух десятилетий.
Липиды являются одними из основных компонентов клеточных мембран. Значительное количество исследований в области физиологии липидов посвящено двум жирным кислотам — ARA и DHA. ARA обнаружена в составе клеточных мембран всех структур организма человека; она является предшественником эйкозаноидов 2-й серии, лейкотриенов 3-й серии и других метаболитов, которые включены в сигнальные системы клеток и процесс генной регуляции. Исследования, посвященные DHA, часто указывают на ее структурную и функциональную роль в составе клеточных мембран.
Эта жирная кислота обнаружена в высокой концентрации в сером веществе головного мозга, а также в палочках и колбочках сетчатки. Исследования постепенного исключения из диеты животных омега-3 жирных кислот показали, что содержащие 22 атома углерода омега-6 ДЦПНЖК (например, 22:5 п—6) способны структурно, но не функционально заменить 22:6 n-3. При неадекватном уровне 22:6 n-3 в тканях выявляются нарушения зрения и познавательных способностей. Было показано, что изменение содержания 22:6 n-3 в тканях влияет на нейротрансмиттерную функцию, активность ионных каналов, сигнальные пути и экспрессию генов.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «физиология человека»
Катаболизм липидов

Внешний обмен липидов
Источник липидов в пище тебе тоже хорошо известен. Это все, что вкусно и вредно. Ну и рыба еще. Расщепление липидов происходит в двенадцатиперстной кишке, но тут уже есть чуть больше нюансов, чем с глюкозой. Липиды очень гидрофобны и собираются, как бабули на скамейке, в мицеллы, обращаясь кнаружи гидрофильными участками молекул. И в этот интимный круг никак не пробиться, если предварительно не эмульгировать их. Для этого в организме придуманы желчные кислоты. Они способствуют распаду липидных капель на миллиарды еще более мелких капель, что делает их доступными для действия панкреатических ферментов [1–3].
В панкреатическом соке, если долго вглядываться, можно найти липазу. Она расщепляет боковые жирные кислоты ТАГ, оставляя после себя 2-моноацилглицерол (2-МАГ).
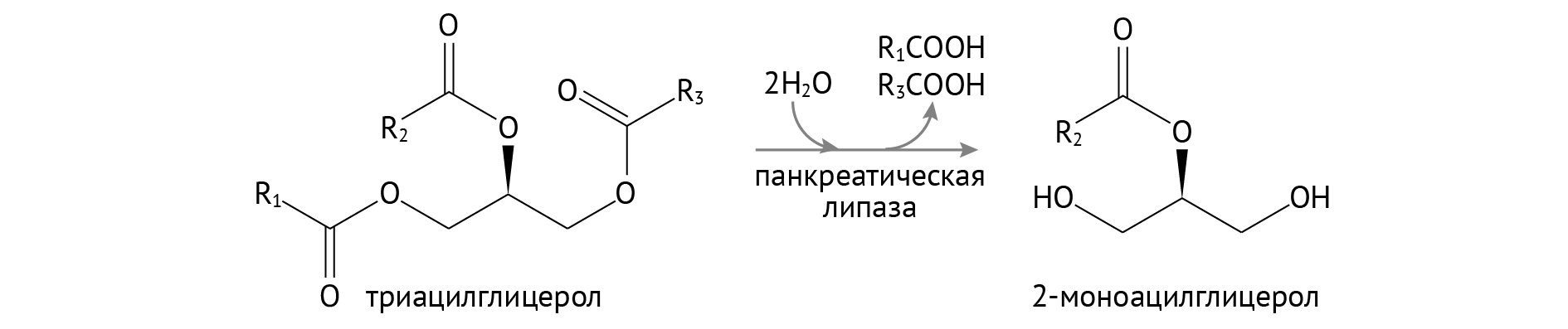
Хотя в ряде случаев все это разрезается до жирных кислот и глицерина.
Продукты расщепления просто превосходно всасываются в энтероциты. Но выбросить жирные кислоты в кровь просто так нельзя. В энтероцитах происходит их ресинтез. Во многом (как ни странно) он повторяет процесс расщепления.
Ресинтез. Чтобы присоединить жирную кислоту к глицерину/2-МАГ, необходимо навести на кислоту марафет. Косметическим элементом для нее служит S-KoA-группа.
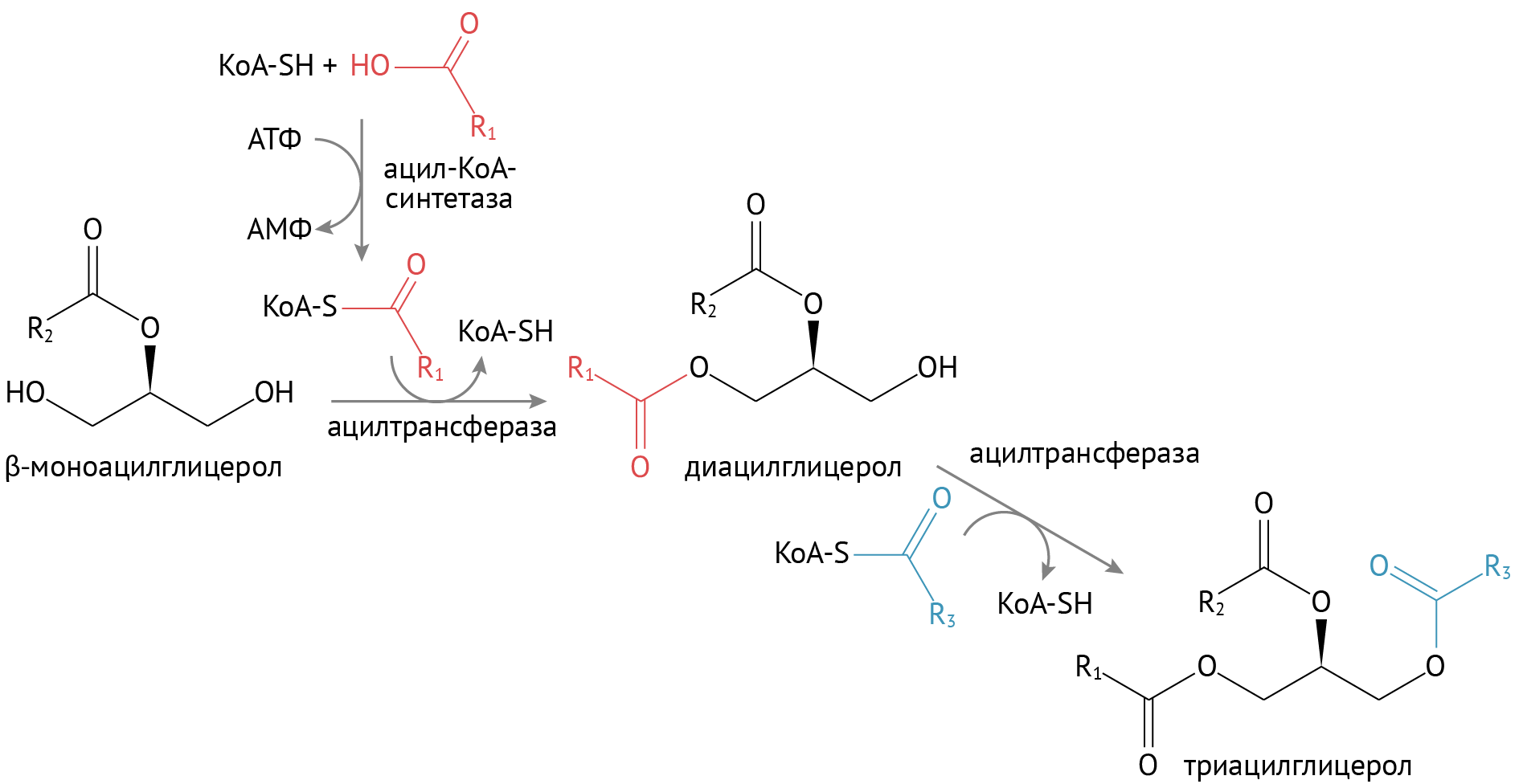
В энтероците собираются еще и фосфолипиды — не менее важные для организма (я бы сказал, самые важные) липиды. Их синтез происходит просто: к диацилглицеролу присоединяется холин.
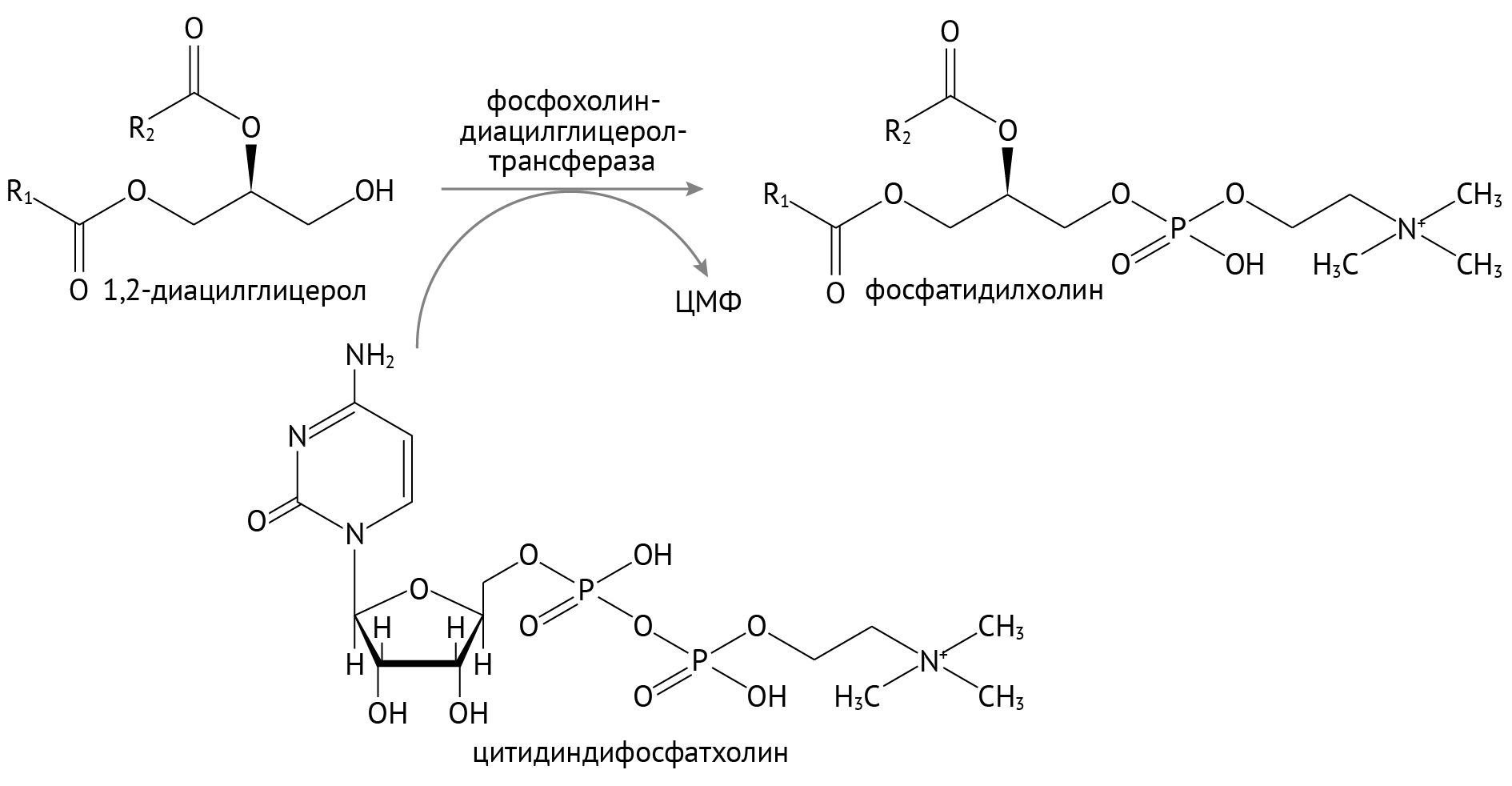
Весь этот комок липидов сбивается в кучу под названием хиломикроны.
Путешествие липидов от кишки до клеток
Липиды, всосавшиеся в кишке, достигают системы верхней полой вены. Но если ты помнишь анатомию, это для тебя не должно быть сюрпризом. Все объясняется просто: хиломикроны всасываются в систему лимфатических капилляров и сосудов. Выбросить их в воротную вену будет насилием над печенью, поэтому мы насилуем лимфатическую дренажную систему. Путь хиломикронов в анатомическом отношении принимает следующий характер. Лимфатические капилляры кишки → лимфатические сосуды брюшной полости → левый грудной лимфатический проток → венозный угол (место слияния левой внутренней яремной и левой подключичной вен, а также место впадения грудного лимфатического протока) → плечеголовная вена → система верхней полой вены [1].
Попадая в систему легочных сосудов, а затем в малый круг, хиломикроны разносятся по тканям. Во многих тканях (особенно в жировой) присутствует фермент липопротеидлипаза. Этот товарищ расщепляет хиломикроны, вычленяя оттуда триглицериды, фосфолипиды, жирные кислоты и вот это вот все. Те благополучно достигают клеток и спокойно попадают внутрь, в цитозоль. Важно отметить, что для активации липопротеинлипазы тканей необходим апопротеин С — II [2].
Хиломикроны приобретают его при взаимодействии с липопротеидами высокой плотности (ЛПВП) в плазме крови.
P. S. Oбмен липопротеидов мы пока опустим и подробно разберем его в другой лекции. А заодно узнаем о биохимических предпосылках атеросклероза — настоящей чумы нашего времени.
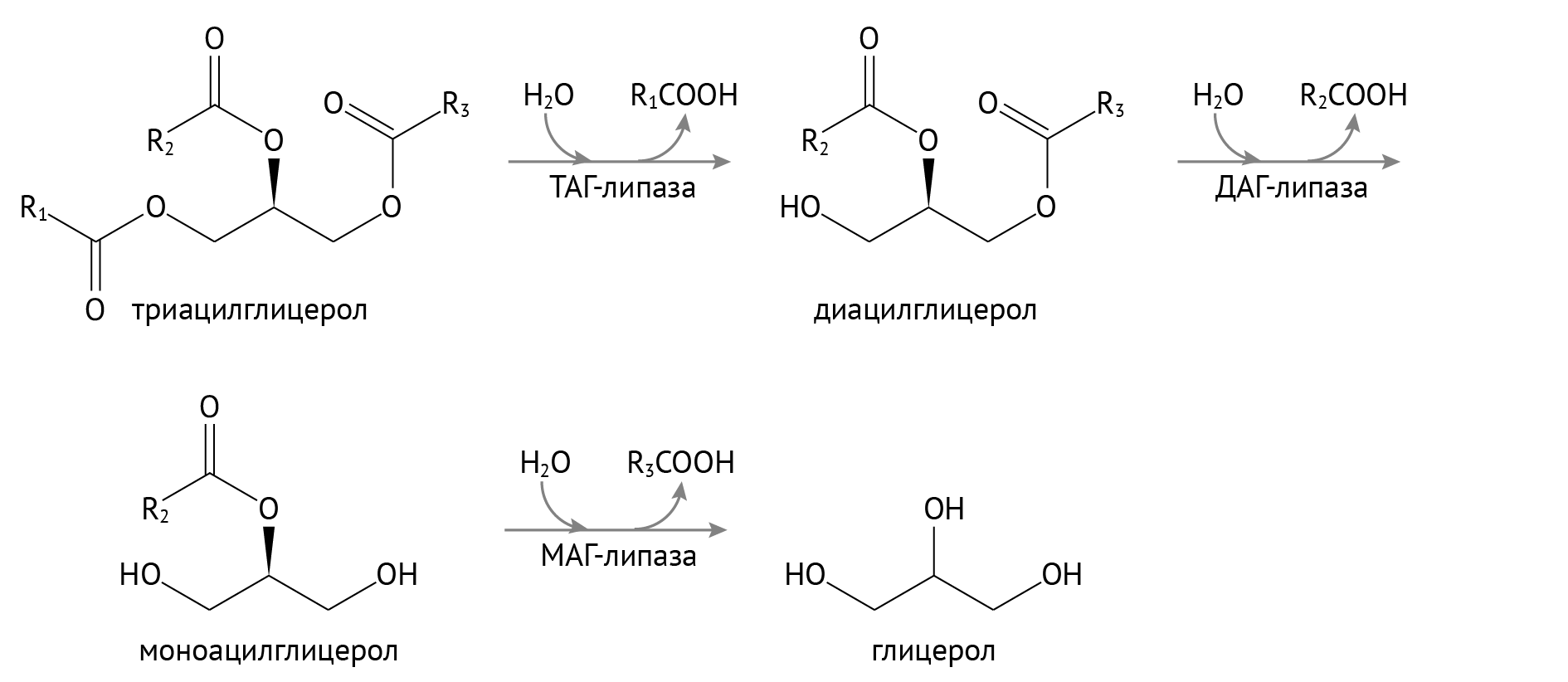
Распад триацилглицеридов в тканях
Триацилглицериды (ТАГ) являются запасной формой энергии в силу того, что при окислении жирных кислот получается просто огромнейшее ее количество. Но чтобы окислить, их нужно достать из лап глицерина.
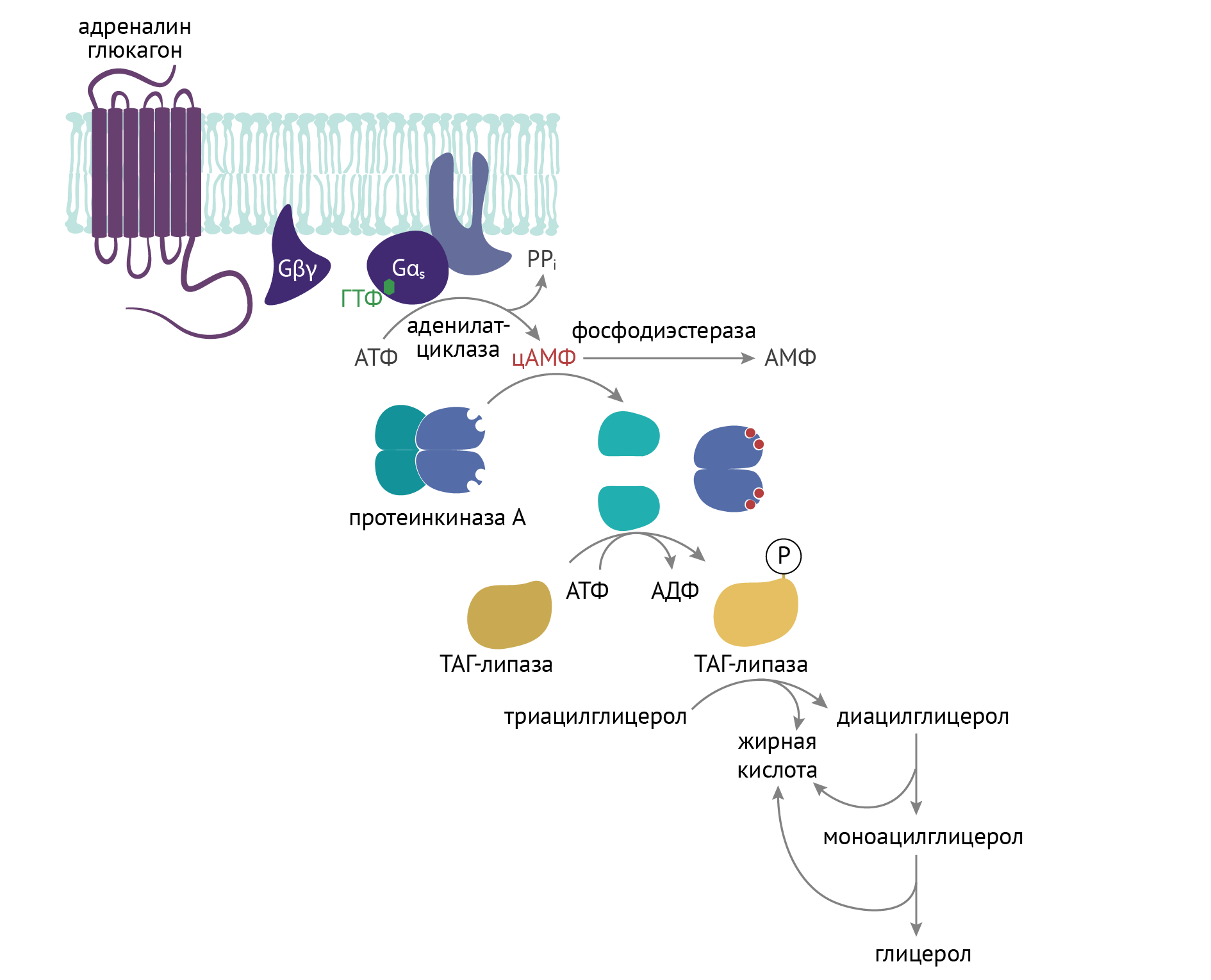
ТАГ-липаза является гормон-чувствительным ферментом, которая очень остро реагирует на критику и на действие контринсулярных гормонов — в первую очередь, адреналина и глюкагона. Он связан с активацией внутриклеточной протеинкиназы А. К слову, с активацией протеинкиназы А сопряжена и активация процесса распада гликогена в печени и мышечной ткани, идущая по аналогичному с активацией ТАГ-липазы механизму [3]:
Не для зубрежки. Литературная минутка
* Большой и неуютный цитозоль. Здесь кипит жизнь, ничего не стоит на месте. Туда-сюда шныряют тРНК, унося с собой несчастную аминокислоту. А вот там можно заметить убиквитинлигазу, которая поймала нарушителя понятий и упаковывает его в протеасомы.
Здесь у каждого есть свое место. Это биологический мегаполис, в водоворот которого случайным образом (хотя что в нашей жизни вообще случайно?) попала жирная кислота. Одинокая, потерянная и несчастная. Слоняясь по клетке и зайдя не в тот угол, она встречается с ацил-КоА-синтетазой. Это встреча не проходит для нее бесследно. Теперь она жирная кислота, но с клеймом в виде S-KoA-группы, и способна вступать в неприличные связи с обитателями цитозоля клетки. Но ей хочется большего, и, как и в макромире, это не всегда приводит к добру…»
Ацил-КоА встречает на своем пути карнитин. Пообщавшись, жирная кислота узнает о существовании чрезвычайно увлекательного места под названием «митохондрия». Будучи легкомысленной и наивной, и в то же время чрезвычайно (в химическом отношении) активной, она недолго ломается и принимает предложение карнитина вписаться в эту удивительную семейную вечеринку.
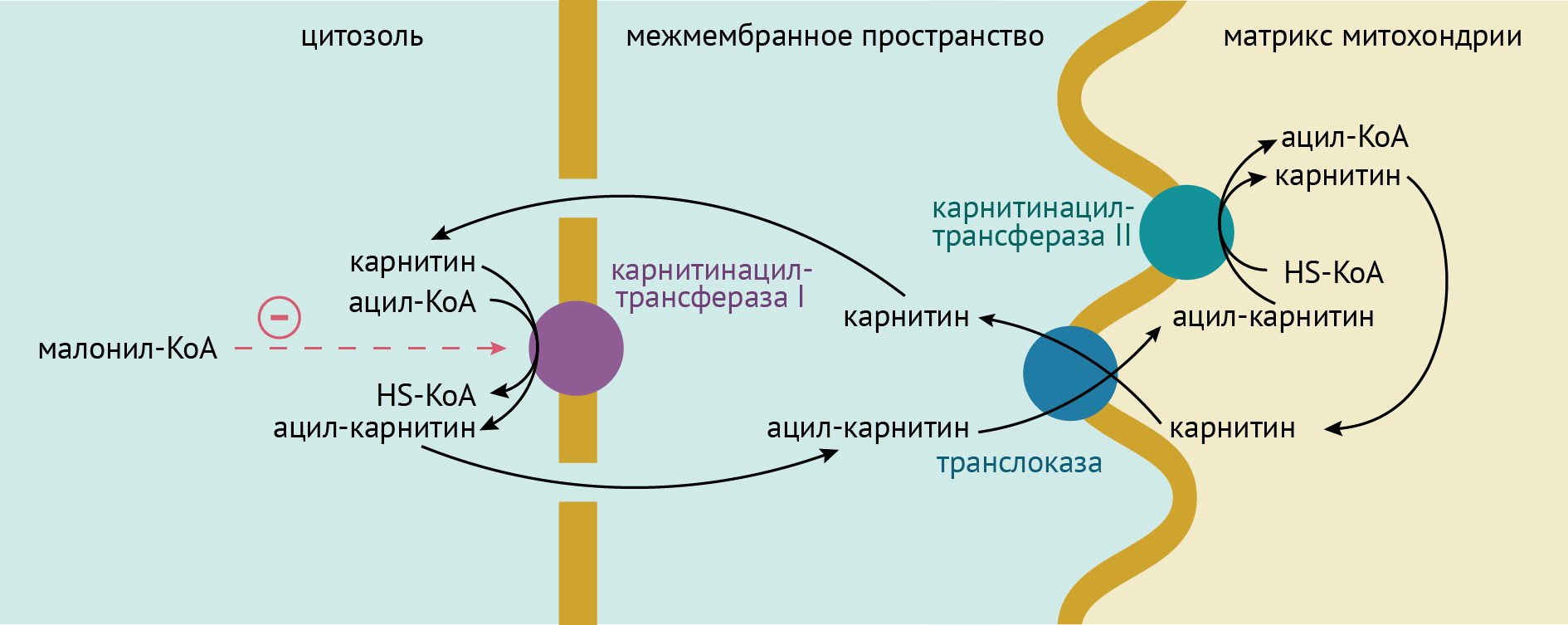
Карнитин — «свой» на этой вечеринке и проносит с собой жирную кислоту, кружась с ней в карнитинацилтрансферазном I танце. Достигая пика любви, карнитин увлекает жирную кислоту через транслоказу в митохондрию. Но на этом их ожидания начинают разниться и карнитин, оставив жирную кислоту с багажом воспоминаний (о котором стыдно рассказывать, но который приятно поворошить), уходит. Преодолев карнитинацилтрансферазу II, ацилкарнитин перестает существовать как пара и распадается на карнитин и ацил-КоА.
β-окисление жирных кислот
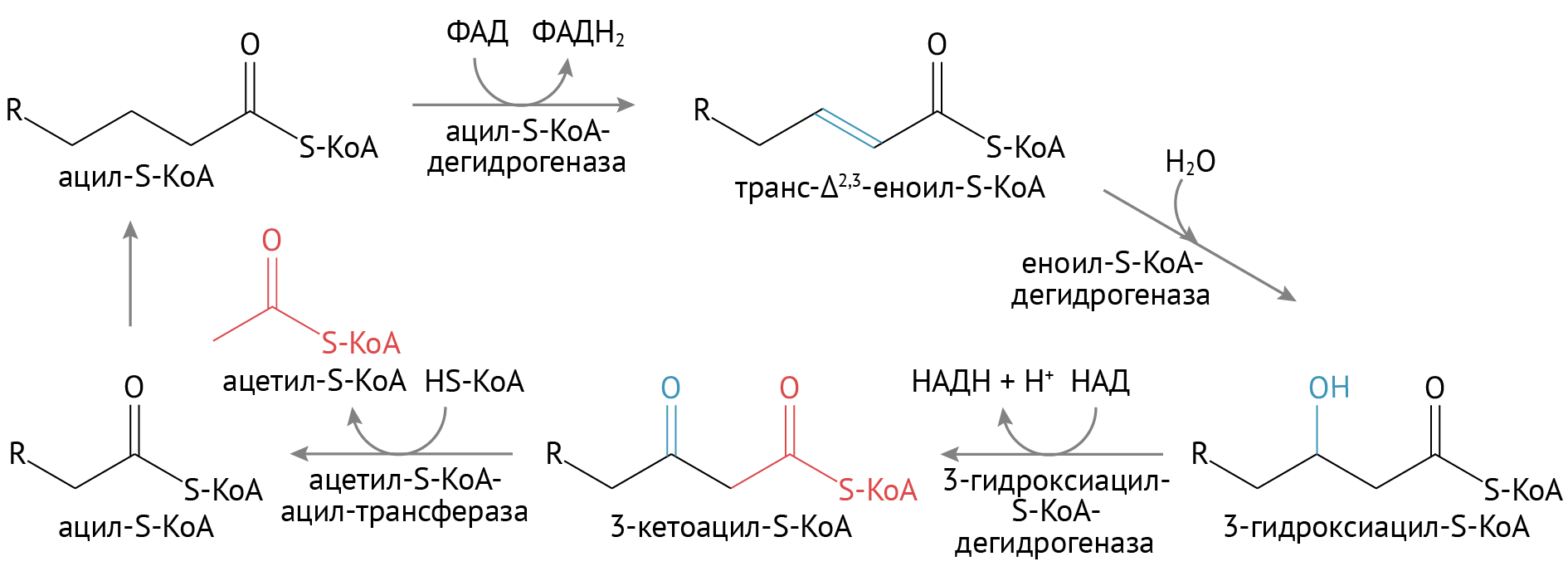
- Сверху представлена схема расщепления насыщенных жирных кислот, т. е. тех кислот, которые исходно не имеют в своей структуре двойных (ненасыщенных) ковалентых связей. Если мы имеем дело с исходно ненасыщенной кислотой, для нее первый цикл начнется со второй реакции.
- Конечный продукт — молекулы ацетил-КоА — необходимый субстрат для цикла трикарбоновых кислот.
- Получение энергии обусловлено восстановлением ФАД до ФАДН2 и НАД до НАДН + Н+. Эти восстановительные эквиваленты играют роль доноров электронов в электрон-транспортной цепи, и, в конечном счете, для получения энергии.
- В целом, окислением одной жирной кислоты можно получить в среднем 90–130 молекул АТФ. Это очень много, особенно для одной молекулы.
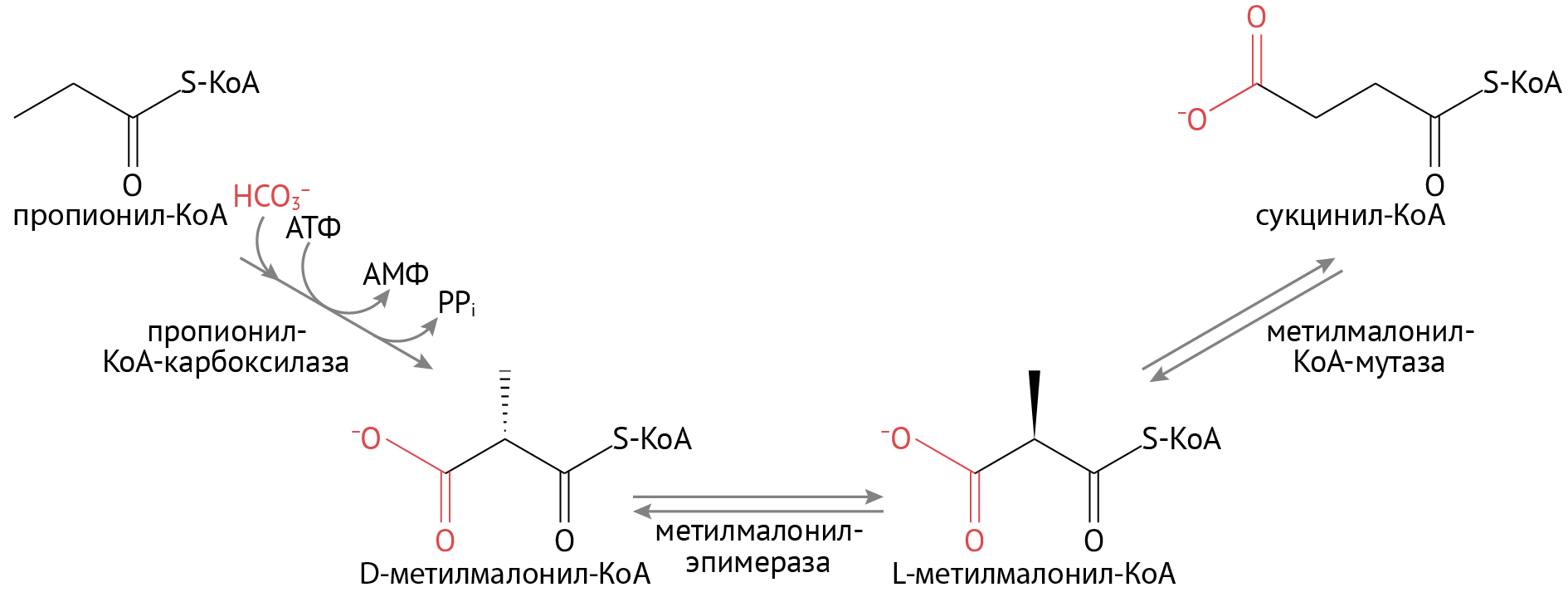
- На последней стадии окисления образуется не ацетил-КоА, а пропионил-КоА. Путем нехитрых превращений она станет сукцинил-КоА — активным участником того же ЦТК.
Метаболизм углеводов и липидов характеризуется наличием еще и сложных анаболических реакций, которыми я пока не хочу нагромождать и без того огромнейшую главу. Какие-то моменты по анаболизму я поясню чуть позже, когда коснусь проблемы заболеваний.
Источники:
- Я. Кольман, К.-Г. Рём. Наглядная биохимия, 5-е издание, 2018 г., с. 154–168.
- Тимин О. А. Основы биологической химии, 2018 г., с. 232–257.
- Портал «Биохимия для студента»:, раздел «Липиды».