
Нуклеопротеиды в кишечнике распадаются на простые белки и нуклеиновые кислоты — РНК и ДНК. В кишечнике под действием специфических нуклеаз, которые вырабатываются в поджелудочной железе, нуклеиновые кислоты расщепляются до мононуклеотидов. Последние подвергаются воздействию фосфатазы, в результате чего от мононуклеотида отщепляется фосфорная кислота. Остаток мононуклеотида — азотистое основание, соединенное с углеводом, носит название нуклеозида — может непосредственно всасываться из кишечника.
Кроме этого, в кишечном соке содержатся активные нуклеазидазы, которые расщепляют нуклеозиды на азотистые основания и пентозы.
Таким образом, в кишечнике происходит распад нуклеопротеидов на исходные составные части, а также мононуклеотиды и нуклеозиды, которые и всасываются в кровь. Кровь доставляет все эти продукты в печень, а также к другим органам и тканям, где они подвергаются специфическим превращениям.
В тканях нуклеиновые кислоты под действием ДНК-азы и РНК-азы распадаются до мононуклеотидов, а последние — на азотистые основания, Н3РО4 и пентозы. Фосфорная кислота и пентозы подвергаются в органах и тканях различным превращениям. Пуриновые основания аденин и гуанин окисляются до мочевой кислоты, которая выводится из организма мочой.

У пиримидиновых оснований в ходе превращений в тканях разрывается кольцо и образуются нециклические соединения. Например, конечными продуктами распада урацила являются β-аланин, один из компонентов коэнзима А (КоА), аммиак и углекислый газ.

Наряду с процессами распада в организме постоянно происходит и синтез нуклеиновых кислот. Необходимые для этого исходные вещества образуются в организме. Пентозы образуются при распаде углеводов (пентозный цикл). Фосфорная кислота всегда присутствует в организме. Синтез пуриновых оснований осуществляется в результате длительных превращений из глицина, глютамина, муравьиной и аспарагиновой кислот и СО2. Пиримидиновые основания образуются из карбамилфосфата, аспарагиновой кислоты и углекислого газа. Под действием ферментов вначале происходит синтез мононуклеотидов, а затем и нуклеиновых кислот РНК и ДНК.
30 И 31. Переваривание нуклеопротеидов в жкт. Распад пуриновых и пиримидиновых нуклеотидов. Подагра.
Нуклеопротеиды – это сложные белки, кот состоят из белковой части и нуклеиновых кислот. В зависимости от содержания нуклеиновых кислот различают ДНП и РНП. Нуклеопротеиды состоят из мононуклеотидов.
Мононуклеотиды выполняют в свободном виде следующие функции:
1) АТФ – универсальный источник энергии 2) УТФ – участвует в синтезе глицерофосфолипидов 3) АМФ – входит в состав НАД и ФАД 4) цАМФ – вторичный посредник в передаче вторичного сигнала.
Распад экзогенных нуклеопротеидов в ЖКТ. Особенности: 1) распад идет гидролитическим способом 2) нуклеотиды и азот. основания, кот получаются в ходе распада практически не используются для синтеза нуклеиновых кислот и кофакторов тканей.
Пуриновые и пиримидиновые азотистые основания всасываются в vena porte и поступают в печень, где происходит их распад (в энтероцитах и гепатоцитах) до конечных продуктов.
Пуриновые азотистые основания окисляются до мочевой кислоты: 1) плохо растворима в Н2О 2) рКм/к= 5,75 при рН меньше 5,75 мочевая кислота в основном находится в протонированной форме, при рН больше 5,75 мочевая кислота образует соли с Na – ураты, кот в 17 раз лучше растворимы в воде, чем мочевая кислота.3) растворимость мочевой кислоты резко снижается при низкой температуре. Роль мочевой кислоты: 1) является конечным продуктом распада 2) проявляет свойства антиоксиданта 3) по структуре похожа на кофеин, поэтому оказывает влияние на функцию цнс. Судьба мочевой кислоты. М/К поступает в кровь, связывается с глобулинами плазмы и это значительно повышает ее растворимость. М/К в крови в норме 0,15-0,42 ммоль/л. из организма м/к в основном выводится с мочой 250-270 миллиграмм в сутки. Повышение концентрации м/к в крови называется гиперурикемия, кот бывает 2 видов: 1) продукционная – образуется в результате повышенного образования м/к 2) ретенционная – в результате снижения выведения с мочой В последствии развивается подагра, при кот наблюдается накопление уратов в тканях, наиболее чувствительны суставы, в кот накопление уратов вызывает воспалительную реакцию и сильные боли, наблюдается образование камней в мочевыводящих путях. Лечение подагры: 1) диета с исключением продуктов с высоким содержанием пуринов ( икра, печень, почки, мясо, красное вино) – болезнь аристократов 2) препарат аллопуринол – структурный аналог гипоксантина, действует как конкурентный ингибитор к , в результате образуется именьше м/к.
Читайте также: Брендовые ткани в рязани
Особенности у новорожденных: на 2-3 день после рождения у детей наблюдается мочекислый инфаркт новорожденных – повышенное выведение м/к с мочой, кот приобретает ярко янтарно-коричневый цвет, связано это с повышенным распадом нуклеопротеидов — физиологическое состояние, кот проходит через 5-7 дней.
Распад пиримидиновых азотистых оснований.
Судьба бета-аланина: 1) трасаминирование и образуется альфа-аланин 2) путь полного окисления 3) идет на синтез карнозина и ансерина – это дипептиды, кот находятся в мышцах и увеличивают амплитуду сокращений в период утомления мышц. Бета-аминоизобутират окиляется до конечных продуктов или в небольшом количестве выводится с мочой, при лейкимии это количество увеличивается за счет повышенного распада клеток.
Катаболизм нуклеопротеидов в тканях организма.
В клетках тканей также происходит катаболизм нуклеопротеидов. В тканях, кроме ДНк-азы и РНК-азы, имеются очень специфические ферменты рестриктазы, которые разрушают определённые, строго специфичные связи в молекулах ДНК и РНК. Эти рестриктазы используются в генной инженерии и биотехнологии («вырезка» и «вставка» генов).
Эндогенные азотистые основания также подвергаются окислению или частично входят в биосинтез.
Пуриновые и пиримидиновые нуклеотиды окисляются по разным метаболическим путям.
а) Катаболизм пуриновых нуклеотидов
Распад аденозина (слайд-плёнка №2) идёт через инозин, т.е. под действием аденозиндезаминазы происходит окислительное дезаминирование аденозина. Недостаточность этого фермента может вызвать тяжёлую форму комбинированного иммунодефицита. Далее, инозин переходит в гипоксантин
(инозиннуклеозидаза), а затем в ксантин. Гипоксантин и ксантин окисляются под действием ксантиоксидазы (КСО) в мочевую кислоту. Фермент КСО содержит молибден (Мо) и способствует пероксидации тканей организма, т.к. в процессе его воздействия на гипоксантин и ксантин накапливается пероксид водорода (Н2О2), что ведёт к перекисному окислению липидов. Образовавшаяся мочевая кислота выделяется в кровь в виде её натриевой соли (ураты), из клеток тканей, а затем фильтруется почечными клубочками, реабсорбируется и снова экскретируется почками. В здоровом организме в крови 3-6 мг/дл мочевой кислоты, а почками выделяются соли мочевой кислоты (ураты) в пределах 0,6-1,2 г/сут (слайд №2). Катаболизм гуанозина осуществляется под действием гуанозиннуклеозидазы, что приводит к образованию гуанина. Затем гуанин подвергается дезаминированию (гуаниндезаминаза) и превращается сразу в ксантин, который с участием КСО переходит в мочевую кислоту. Биологическая роль уратов велика, они могут функционировать как антиоксиданты. Предполагается, что эндогенные антиоксиданты – ураты – заменяют у приматов аскорбиновую кислоту (вит. С), способность к синтезу которой у них утрачена. Известно, что гулонолактон не восстанавливается в аскорбат из-за потери фермента
Накопление уратов в организме вызывает заболевание подагра, которая проявляется избыточным отложением солей мочевой кислоты в суставах, суставных сумках и т.д. Известно, что конечные продукты катаболизма пуринов (мочевая кислота и её натриевая соль) обладают слабой растворимостью. Лечебным препаратом, применяемым для торможения синтеза уратов, является аллопуринол, механизм которого сводится к конкурентному торможению фермента ксантиоксидазы.
Причины гиперурикемии – повышение концентрации уратов в крови различны: во-первых, алиментарное происхождение; далее, усиленный распад ядерного вещества клеток (лейкоз, злокачественная опухоль, радиотерапия); затем – нарушение выделительной функции почек.
б) Катаболизм пиримидиновых нуклеотидов.
Известно, что к пиримидиновым нуклеотидам относят уридиловую, цитиловую и тимидиловую кислоты. В результате катаболизма пиримидиновых нуклеотидов образуются хорошо растворимые конечные продукты.
Катаболизм цитидина протекает в печени (преимущественно) через реакцию окислительного дезаминирования в уридин, который превращается в урацил (уридиннуклеозидаза), а затем через образование мочевины (СО2 и NН3) и бета-аланина (слайд-плёнка №3). Последний, либо выделяется через почки с мочой, или попадает в мышцы и является предшественником эндогенных антиоксидантов – ансерина и карнозина. Катаболизм тимидина проходит стадию образования тимина (тимидин-нуклеозидазы), а затем через образование мочевины (СО2 и NН3) и бета-аминоизобутират, который выводится почками. Экскреция бета-аминоизобутирата у человека увеличивается при лучевой болезни, при лейкемии и отражает ускорение гибели клеток и деструкции ДНК.
Читайте также: Как правильно резать ткань для коврика
Тема 5. Обмен нуклеопротеинов. Биосинтез и распад пуриновых и пиримидиновых нуклеотидов.
Практическая значимость темы. Нуклеотиды — одни из наиболее важных компонентов клетки, принимающие участие во многих биохимических процессах. Они являются структурными единицами ДНК и РНК, участвуют в активации промежуточных продуктов реакций биосинтеза (УДФ-глюкоза, ЦДФ-холин), входят в состав коферментов (НАД, ФАД, КоА-SH, АТФ), служат аллостерическими регуляторами процессов метаболизма (АМФ) и внутриклеточными посредниками действия гормонов (цАМФ, цГМФ). Образование нуклеотидов в организме происходит главным образом в процессе поэтапной сборки из эндогенных предшественников, реже путём реутилизации азотистых оснований, высвобождаемых при расщеплении нуклеиновых кислот в тканях. Скорость синтеза пуриновых и пиримидиновых рибо- и дезоксирибонуклеотидов в организме является объектом тонкой регулировки. Знание процессов биосинтеза и распада нуклеотидов позволяет понять патогенез ряда заболеваний, связанных с нарушениями обмена пуринов и пиримидинов (подагра, ксантинурия, оротацидурия), грамотно использовать в качестве противоопухолевых препаратов структурные аналоги азотистых оснований, нуклеозидов (6-меркаптопурин, 5-фторурацил, фторафур) и антагонисты коферментов, участвующих в биосинтезе нуклеотидов (аметоптерин, аминоптерин).
Цель занятия. После изучения данной темы студент должен знать особенности биосинтеза и распада пуриновых и пиримидиновых нуклеотидов, механизмы регуляции этих процессов и основные виды их нарушений, уметь применять приобретённые знания для решения теоретических и практических задач.
Исходный уровень знаний.
Строение пуриновых и пиримидиновых азотистых оснований, нуклеозидов, нуклеотидов, их роль в организме.
Строение аминокислот (глицин, глутамин, аспартат).
Пентозофосфатный путь окисления: роль в организме.
Коферментные функции витаминов (фолиевая кислота).
Механизмы регуляции активности ферментов.
Принципы диагностики врождённых дефектов ферментов.
5.1. Катаболизм нуклеопротеинов.
5.1.1. Нуклеопротеины – сложные белки, содержащие в качестве простетической группы нуклеиновые кислоты (РНК или ДНК). В зависимости от того, какая кислота входит в их состав, различают рибонуклеопротеины и дезоксирибонуклеопротеины.
Структурными единицами РНК и ДНК являются нуклеотиды, каждый из которых в свою очередь состоит из азотистого основания и пентозы (образующих нуклеозид), соединённых с остатком фосфорной кислоты.
Пентоза и азотистое основание в каждом нуклеотиде связаны N-гликозидной связью, фосфат и пентоза – сложноэфирной связью. Отдельные нуклеотиды в ДНК или РНК связаны 3’,5’-фосфодиэфирной связью (см. рисунок).
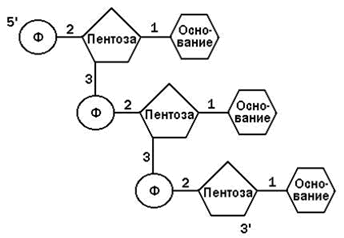
Рисунок 5.1. Схема строения фрагмента полинуклеотидной цепи. Примечание. Пентоза – рибоза в РНК, дезоксирибоза в ДНК. Азотистое основание – аденин, гуанин, цитозин, урацил в РНК, аденин, гуанин, цитозин, тимин в ДНК. Цифрами обозначены типы связей: 1 – N-гликозидные; 2 – сложноэфирные в нуклеотиде; 3 – 3’,5’-фосфодиэфирные (между нуклеотидами).
5.1.2. Катаболизм нуклеопротеинов в желудочно-кишечном тракте. Нуклеопротеины, поступающие в организм с пищей, подвергаются расщеплению под действием протеолитических ферментов кишечника. Высвобождаемые нуклеиновые кислоты гидролизуются при участии рибонуклеаз и дезоксирибонуклеаз панкреатического сока до мононуклеотидов. Действие панкреатических нуклеаз дополняют фосфоэстеразы кишечного сока. Схематично это можно представить следующим образом:
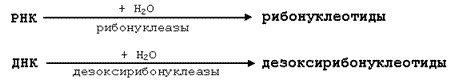
Далее, под воздействием нуклеотидаз и фосфатаз происходит гидролиз нуклеотидов до нуклеозидов, которые либо всасываются, либо под воздействием ферментов слизистой кишечника расщепляются до пуриновых и пиримидиновых оснований.
В организме человека большая часть пуриновых и пиримидиновых оснований, высвободившихся из нуклеиновых кислот, которые поступают с пищей, превращается в конечные продукты (при этом не происходит их включения во вновь образующиеся молекулы нуклеиновых кислот в тканях организма). То есть, нуклеиновые кислоты пищи практически не выступают в роли поставщика непосредственных предшественников нуклеиновых кислот тканей организма.
В то же время нуклеотиды и нуклеозиды, введённые парентерально, могут включаться в нуклеиновые кислоты без всяких изменений. Это послужило основой методов исследования метаболизма нуклеиновых кислот путём введения меченных радиоактивными изотопами азотистых оснований.
Читайте также: Ткань для 21 волги
46. Распад нуклеиновых кислот, нуклеазы пищеварительного тракта и тканей.
Нуклеиновые кислоты поступают в организм с пищей главным образом в составе нуклеопротеинов и высвобождаются в результате действия протеолитических ферментов желудочно-кишечного тракта. Далее под действием дезоксирибонуклеазы и рибонуклеазы панкреатического сока нуклеиновые кислоты гидролизуются до нуклеотидов. Нуклеотиды под воздействием нуклеотидаз или фосфатаз распадаются до нуклеозидов, которые могут всасываться или гидролизоваться далее до азотистых оснований и пентоз. В тканях нуклеиновые кислоты гидролизуются дезоксирибонуклеазами (ДНК-азы) и рибонуклеазами (РНК-азы) до нуклеотидов, которые под действием нуклеотидаз теряют остаток фосфора. Образующиеся нуклеозиды пуринового и пиримидинового ряда подвергаются дальнейшему катаболизму.

47. Распад пуриновых нуклеотидов.
48. Биосинтез пуриновых нуклеотидов, происхождение атомов «С» и «N» в пуриновом кольце.
Происхождение атомов C and N. Тут мы таки видим как они образуются

Синтез не помещается, да и он же не требуется как в учебнике, сойдет же так как на лекции. А я не умею рисовать формулы и схемы в ворде. Поэтому вот вам поле рисования
49. Инозиновая кислота как предшественник пуриновых мононуклеотидов
50. Распад пиримидиновых нуклеотидов.

51. Биосинтез пиримидиновых нуклеотидов.
Фонд пирймидиновых нуклеотидов, подобно пуриновым нуклеотидам, в основном синтезируется из простых предшественников de novo, и только 10-20% от общего количества образуется по «запасным» путям из азотистых оснований или нуклеозидов.
Образование пиримидиновых нуклеотидов DE NOVO
Пиримидиновое кольцо синтезируется из простых предшественников: глутамина, СО2 и аспарагиновой кислоты и затем связывается с рибозо-5-фосфатом, полученным от ФРДФ.
Процесс протекает в цитозоле клеток. Синтез ключевого пиримидинового нуклеотида — УМФ идёт с участием 3 ферментов, 2 из которых полифункциональны.
У млекопитающих ключевой, регуляторной реакцией в синтезе пирймидиновых нуклеотидов является синтез карбамоилфосфата из глутамина, СО2 и АТФ, в реакции катализируемой кар-бамоилфосфатсинтетазой II (КФС II), которая протекает в цитозоле клеток (рис. 10-12). В реакции NH2-гpyппa карбамоилфосфата образуется за счёт амидной группы глутамина.

Рис. 10-12. Синтез карбамоилфосфата.
Карбамоилфосфат, использующийся на образование пирймидиновых нуклеотидов, является продуктом полифункционального фермента, который наряду с активностью КФС II содержит каталитические центры аспартаттранскарбамоилазы и дигидрооротазы. Этот фермент назвали «КАД-фермент». Объединение первых трёх ферментов метаболического пути в единый полифункциональный комплекс позволяет использовать почти весь синтезированный в первой реакции карбамоилфос-фат на взаимодействие с аспартатом и образование карбамоиласпартата, от которого отщепляется вода и образуется циклический продукт — дигидрооротат (рис. 10-13).

Рис. 10-13. Биосинтез УМФ de novo.
Отщепляясь от КАД-фермента, дигидрооротат подвергается дегидрированию NAD-зависимой дигидрооротатдегидрогеназой и превращается в свободное пиримидиновое основание — оротовую кислоту, или оротат.
В цитозоле оротат становится субстратом бифункционального фермента — УМФ-синтазы, которая обнаруживает оротатфосфорибозилтранс-феразную и ОМФ-декарбоксилазную активности. Первоначально фосфорибозильный остаток от ФРДФ переносится на оротат и образуется нук-леотид — оротидин-5′-монофосфат (ОМФ), декарбоксилирование которого даёт уридин-5-монофосфат (УМФ).
Таким образом, шесть последовательных реакций синтеза пиримидиновых нуклеотидов осуществляются тремя ферментами, которые кодируются в геноме человека тремя различными структурными генами.
Биосинтез УДФ, УТФ и иитидиловых нуклеотидов
УМФ под действием специфических нуклео-зидмонофосфат (НМФ) и нуклеозиддифосфат (НДФ) киназ превращается в УДФ и УТФ в результате переноса γ-фосфатного остатка АТФ на соответствующий субстрат.
НМФ-киназа катализирует следующую реакцию:
ЦТФ синтетаза катализирует амидирование УТФ (рис, 10-14), осуществляя АТФ-зависимое замещение кетогругшы урацила на амидную группу глутамина с образованием цитидин-5′-трифосфата (ЦТФ).
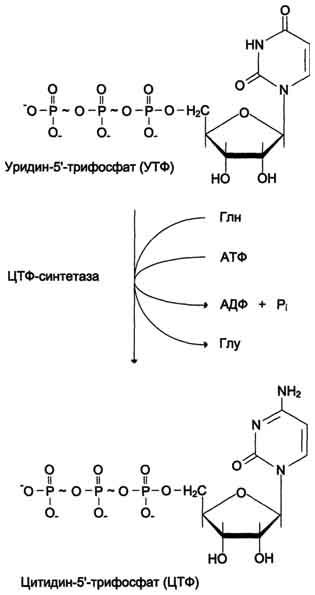
Рис. 10-14. Синтез ЦТФ из УТФ.
«Запасные» пути синтеза пиримидиновых нуклеотидов
Использование пиримидиновых оснований и нуклеозидов в реакциях реутилизации препятствует катаболизму этих соединений до конечных продуктов с расщеплением пиримидинового кольца. В ресинтезе пиримидинов участвуют некоторые ферменты катаболизма нуклеотидов. Так, уридинфосфорилаза в обратимой реакции может рибозилироватъ урацил с образованием уридина.
Урацил + Рибозо-1-фосфат → Уридин + Н3РО4.
Превращение нуклеозидов в нуклеотиды катализирует уридин-цитидинкиназа.
Часть ЦМФ может превращаться в УМФ под действием цитидиндезаминазы и пополнять запасы уридиловых нуклеотидов.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом