
ПИРИМИДИНОВЫЙ ОБМЕН — совокупность реакций синтеза и распада пиримидиновых нуклеотидов, к-рые наряду с их пуриновыми аналогами используются в живых организмах для синтеза нуклеиновых кислот. П. о. особенно интенсивно совершается в растущих тканях. Ингибирование каких-либо стадий П. о. может привести к замедлению и даже полной остановке деления и роста клеток. Генетически обусловленное нарушение П. о. может быть причиной наследственных заболеваний, напр, оротацидурии (см.).
Интенсивное изучение биосинтеза пиримидиновых оснований (см.) — урацила, тимина и цитозина — началось в 50-е гг. 20 в., когда благодаря работам Лагерквиста (L. Lagerkvist) и Рейнхарда (P. Reinhard), a также других исследователей было установлено, что промежуточным продуктом биосинтеза пиримидиновых нуклеотидов является оротовая кислота (см.). Одновременно было обнаружено, что предшественником одного из атомов азота в пиримидиновом кольце является аммиак, а один из атомов углерода включается в пиримидиновое кольцо из двуокиси углерода. Остальные углеродные атомы пиримидинового кольца принадлежат аспарагиновой к-те.

Биосинтетические реакции П. о. можно разделить на следующие стадии: образование пиримидинового кольца в форме оротовой к-ты; превращение оротовой к-ты в уридиловую к-ту (уридин-5-монофосфат); преобразование уридиловой к-ты, катализируемое соответствующими киназами, в пиримидиновые нуклеотиды, используемые для синтеза РНК (рибонуклеотиды), ДНК (дезоксирибонуклеотиды) и ряда других соединений. Исходными веществами для синтеза пиримидиновых нуклеотидов являются аспарагиновая к-та и карбамоилфосфат, который в свою очередь синтезируется в организме из двуокиси углерода и аммиака при участии АТФ (рис. 1).
Конечные продукты цепи биосинтетических реакций П. о.— уридин-трифосфат (УТФ) и цитидинтрифос-фат (ЦТФ) — используются для синтеза РНК. Для синтеза ДНК необходимо образование дезоксирибо-нуклеотидов: дезоксицитидинтрифос-фата (дЦТФ) и дезокситимидинтри-фосфата (дТТФ). Цитидиндифосфат (ЦДФ), образующийся под действием фосфатазы из ЦТФ, превращается в дЦДФ путем прямого восстановления рибозного кольца, дТМФ получается путем метилирования пиримидинового кольца, входящего в дУМФ.

Распад пиримидиновых нуклеотидов начинается с отщепления от них о-фосфорной к-ты под действием соответствующих нуклеотидаз. Образовавшиеся нуклеозиды подвергаются фосфоролизу с освобождением свободных пиримидиновых оснований и рибозо- или дезоксирибозофосфатов. Свободные цитозин и 5-метилцитозин сначала дезаминируются. Катаболические превращения урацила и тимина заключаются в восстановлении пиримидинов в дигидропроизводные, размыкании кольца, ведущем к образованию уреидокислот. На следующих стадиях происходит удаление аммиака и двуокиси углерода, в результате чего образуется бета-аланин или его метилированное производное — бета-аминоизомасляная к-та; бета-аланин может подвергаться дальнейшему распаду (рис. 2).
Необходимо отметить, что синтез нуклеиновых кислот (см.) регулируется не столько на стадии полимеризации нуклеотидов, сколько на стадии их синтеза. Образование пиримидиновых нуклеотидов регулируется по механизму обратной связи. Один из конечных продуктов цепи биосинтетических реакций пиримидинового обмена — ЦТФ путем прямого аллостерического воздействия на первый специфический фермент синтеза пиримидиновых нуклеотидов аспартат-карбамоилтрансферазу ингибирует реакцию синтеза карбамоиласпартата. Пиримидиновые нуклеотиды также ингибируют синтез трех ферментов П. о.: аспартат-карбамоилтрансферазы, ди-гидрооротазы и оротатредуктазы. Еще одним регуляторным ферментом является киназа, катализирующая фосфорилирование тимидиновых нуклеотидов (превращение дТМФ в дТДФ и дТТФ). Активность этого фермента очень низка в тканях, состоящих из неделящихся клеток, напр, в ткани печени, однако она резко увеличивается сразу же после частичной гепатэктомии. После частичной гепатэктомии возрастает и активность аспартат-карбамоилтрансферазы. По мере восстановления нормального размера органа активность этих ферментов возвращается к норме. Увеличение активности аспартат-карбамоилтрансферазы наблюдают также при гепатоме Новикова. Необходимо отметить, что во всех быстрорастущих тканях, где идет интенсивный синтез нуклеиновых к-т, практически отсутствуют ферменты, участвующие в процессе распада пиримидиновых нуклеотидов.
К числу заболеваний, при которых нарушается синтез пиримидиновых нуклеотидов, следует отнести злокачественную анемию (см. Пернициозная анемия). Леч. действие витамина B12 (см. Цианокобаламин) и фолиевой кислоты (см.) при этом заболевании связано, по-видимому г с участием этих витаминов в синтезе тимина, а также других пиримидинов и пуринов.
Библиография: Дэвидсон Дж. Н. Биохимия нуклеиновых кислот, пер. с англ., М., 1976; Кросби Дж. Биосинтез, пиримидиннуклеотидов, в кн.; Нуклеиновые кислоты, под ред. Э. Чаргаффа и Дж. Дэвидсона, пер. с англ., с. 270, М.„ 1962; Ленинджер А. Биохимия, пер. с англ., М., 1976.
2.2. Распад пуриновых и пиримидиновых нуклеотидов до конечных продуктов в тканях.
Распад пуриновых нуклеотидов.
Аденозин и гуанозин, которые образуются при гидролизе пуриновых нуклеотидов, подвергаются ферментативному распаду с образованием конечного продукта – мочевой кислоты, которая выводится с мочой из организма.
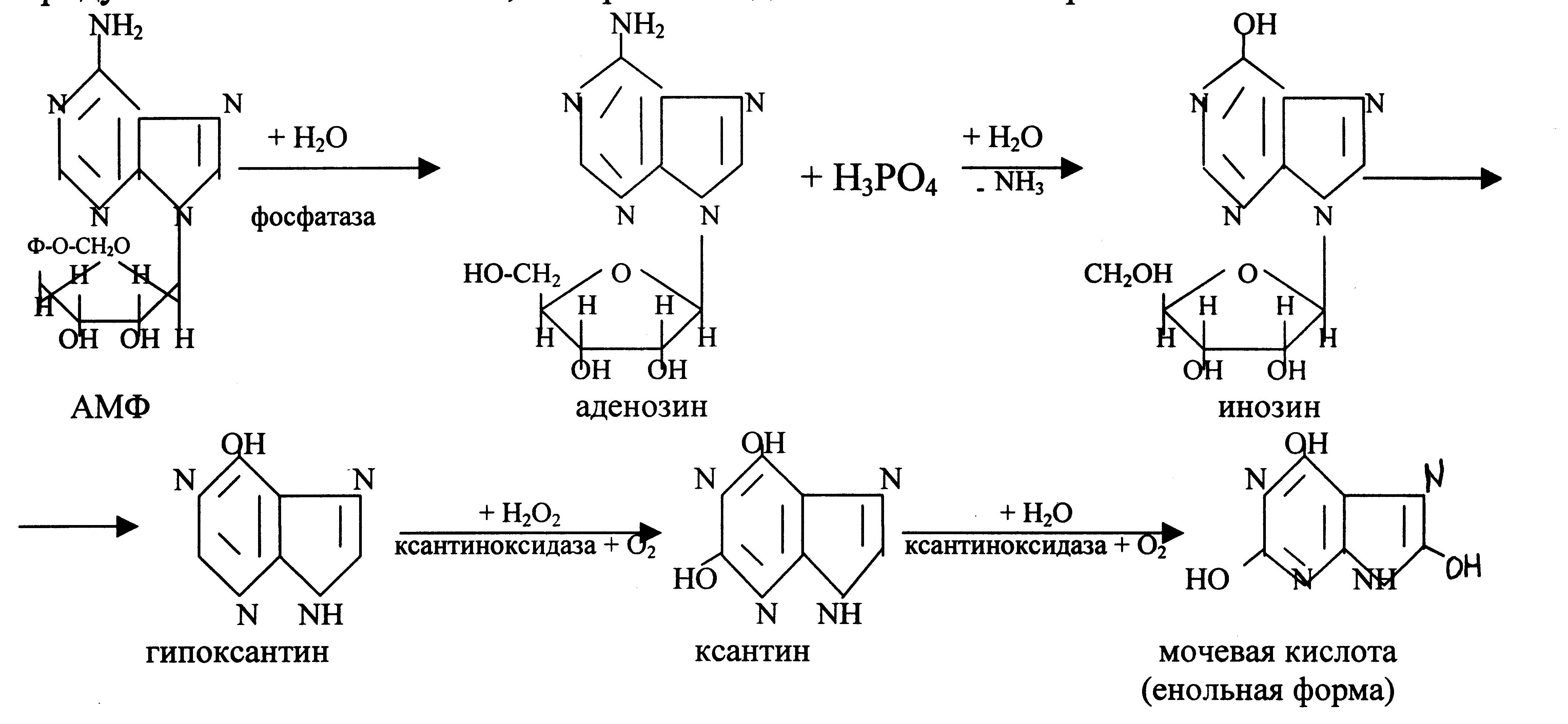
Распад пиримидиновых нуклеотидов.
Начальные этапы этого процесса катализируются специфическими ферментами. Конечные продукты: СО2, NН3, мочевина, β-аланин, β-аминоизомасляная кислота. β-аланин используется для синтеза дипептидов мышц – карнозина и ансерина или выделяется с мочой.

Биосинтез пуриновых, пиримидиновых нуклеотидов в тканях.
Биосинтез пуриновых мононуклеотидов.
Первоначальным соединением синтеза служит Д-рибозо-5-фосфат, который является продуктом пентозофосфатного цикла и на который переносится пирофосфатная группа АТФ. Образовавшийся 5-фосфорибозил-1-пирофосфат (ФРПФ) взаимодействует с глутамином, который является донором NН2-группы в результате чего образуется β-5-фосфорибозил-амин. Эта стадия становится ключевой в синтезе пуринов. Затем присоединяется молекула глицина к свободной NН2-группе β-5-фосфорибозил-амина с образованием глицинамидрибонуклеотида. Еще через несколько стадий образуется первый пуриновый нуклеотид инозинмонофосфат (ИМФ), из которого затем синтезируются остальные нуклеозидфосфаты.

Биосинтез пиримидиновых нуклеотидов
Первоначальными соединениями этого процесса являются карбамоилфосфат и аспарагиновая кислота. Из них через длинную цепь реакций образуется уридинмонофосфат (УМФ) и остальные пиримидиновые нуклеотиды.

2.4. Заболевания, связанные с нарушением обмена нуклеотидов: подагра, синдром Леша-Нихена.
Гиперурикемия – повышение в плазме крови концентрации мочевой кислоты. Вследствие гиперурикемии может развиться подагра.
Подагра – заболевание, вызванное нарушением обмена нуклеиновых кислот. В хрящах, сухожилиях, в суставных сумках, иногда в почках, коже, мышцах откладываются кристаллы мочевой кислоты и уратов. Вокруг этих отложений образуется воспаление и грануляционный вал, который окружает омертвевшую ткань, при этом образуются подагрические узлы — тофусы (в суставах пальцев рук, ног, в хрящах ушной раковины), что сопровождается деформацией и болезненностью пораженных суставов. К характерным признакам подагры относятся повторяющиеся приступы острого воспаления суставов (чаще всего мелких) – острого подагрического артрита. Обычно больные склонны к атеросклерозу и гипертонии. В их крови наблюдается большая концентрация мочевой кислоты – гиперурикемия. В течение нескольких дней перед приступом подагры увеличивается выделение воды и хлорида натрия с мочой, т.е. сдвигается водно-солевой баланс. Вследствие этого возрастает концентрация мочевой кислоты в крови и отложение ее в тканях. Как правило, подагра генетически детерминирована и носит семейный характер. Она вызвана нарушениями в работе фосфорибозилдифосфата (ФРДФ) синтетазы или гипоксантингуанин- или аденинфосфорибозилтрансфераз. К другим характерным проявлениям относят нефропатию, при которой наблюдают образование уратных камней в мочевыводящих путях.
Синдром Леша-Нихена – тяжелая форма гиперурикемии, которая наследуется как рецессивный признак, сцепленный с Х-хромосомой. Проявляется только у мальчиков. Кроме симптомов подагры наблюдаются церебральные параличи, нарушение интеллекта, попытки наносить себе раны (укусы губ, пальцев). Связана болезнь с дефектом фермента гипоксантин-гуанин-фосфорибозилтрансферазы, которая катализирует превращение гипоксантина и гуанина в гуанинимонофосфат (ГМФ), поэтому они превращаются в мочевую кислоту. В первые месяцы жизни неврологические расстройства не обнаруживаются, но на пеленках отмечают розовые пятна, вызванные присутствием в моче кристаллов мочевой кислоты. При отсутствии лечения больные погибают в возрасте до 10 лет из-за нарушения функции почек.
Читайте также: Хрящевая соединительная ткань местонахождение
Основной препарат для лечения гиперурикемии – аллопуринол (структурный аналог гипоксантина).
Тема 5. Обмен нуклеопротеинов. Биосинтез и распад пуриновых и пиримидиновых нуклеотидов.
Практическая значимость темы. Нуклеотиды — одни из наиболее важных компонентов клетки, принимающие участие во многих биохимических процессах. Они являются структурными единицами ДНК и РНК, участвуют в активации промежуточных продуктов реакций биосинтеза (УДФ-глюкоза, ЦДФ-холин), входят в состав коферментов (НАД, ФАД, КоА-SH, АТФ), служат аллостерическими регуляторами процессов метаболизма (АМФ) и внутриклеточными посредниками действия гормонов (цАМФ, цГМФ). Образование нуклеотидов в организме происходит главным образом в процессе поэтапной сборки из эндогенных предшественников, реже путём реутилизации азотистых оснований, высвобождаемых при расщеплении нуклеиновых кислот в тканях. Скорость синтеза пуриновых и пиримидиновых рибо- и дезоксирибонуклеотидов в организме является объектом тонкой регулировки. Знание процессов биосинтеза и распада нуклеотидов позволяет понять патогенез ряда заболеваний, связанных с нарушениями обмена пуринов и пиримидинов (подагра, ксантинурия, оротацидурия), грамотно использовать в качестве противоопухолевых препаратов структурные аналоги азотистых оснований, нуклеозидов (6-меркаптопурин, 5-фторурацил, фторафур) и антагонисты коферментов, участвующих в биосинтезе нуклеотидов (аметоптерин, аминоптерин).
Цель занятия. После изучения данной темы студент должен знать особенности биосинтеза и распада пуриновых и пиримидиновых нуклеотидов, механизмы регуляции этих процессов и основные виды их нарушений, уметь применять приобретённые знания для решения теоретических и практических задач.
Исходный уровень знаний.
Строение пуриновых и пиримидиновых азотистых оснований, нуклеозидов, нуклеотидов, их роль в организме.
Строение аминокислот (глицин, глутамин, аспартат).
Пентозофосфатный путь окисления: роль в организме.
Коферментные функции витаминов (фолиевая кислота).
Механизмы регуляции активности ферментов.
Принципы диагностики врождённых дефектов ферментов.
5.1. Катаболизм нуклеопротеинов.
5.1.1. Нуклеопротеины – сложные белки, содержащие в качестве простетической группы нуклеиновые кислоты (РНК или ДНК). В зависимости от того, какая кислота входит в их состав, различают рибонуклеопротеины и дезоксирибонуклеопротеины.
Структурными единицами РНК и ДНК являются нуклеотиды, каждый из которых в свою очередь состоит из азотистого основания и пентозы (образующих нуклеозид), соединённых с остатком фосфорной кислоты.
Пентоза и азотистое основание в каждом нуклеотиде связаны N-гликозидной связью, фосфат и пентоза – сложноэфирной связью. Отдельные нуклеотиды в ДНК или РНК связаны 3’,5’-фосфодиэфирной связью (см. рисунок).
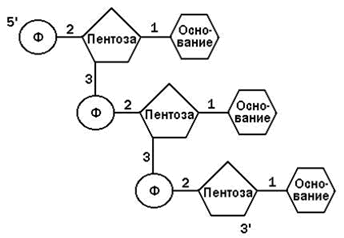
Рисунок 5.1. Схема строения фрагмента полинуклеотидной цепи. Примечание. Пентоза – рибоза в РНК, дезоксирибоза в ДНК. Азотистое основание – аденин, гуанин, цитозин, урацил в РНК, аденин, гуанин, цитозин, тимин в ДНК. Цифрами обозначены типы связей: 1 – N-гликозидные; 2 – сложноэфирные в нуклеотиде; 3 – 3’,5’-фосфодиэфирные (между нуклеотидами).
5.1.2. Катаболизм нуклеопротеинов в желудочно-кишечном тракте. Нуклеопротеины, поступающие в организм с пищей, подвергаются расщеплению под действием протеолитических ферментов кишечника. Высвобождаемые нуклеиновые кислоты гидролизуются при участии рибонуклеаз и дезоксирибонуклеаз панкреатического сока до мононуклеотидов. Действие панкреатических нуклеаз дополняют фосфоэстеразы кишечного сока. Схематично это можно представить следующим образом:
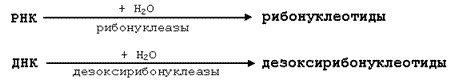
Далее, под воздействием нуклеотидаз и фосфатаз происходит гидролиз нуклеотидов до нуклеозидов, которые либо всасываются, либо под воздействием ферментов слизистой кишечника расщепляются до пуриновых и пиримидиновых оснований.
В организме человека большая часть пуриновых и пиримидиновых оснований, высвободившихся из нуклеиновых кислот, которые поступают с пищей, превращается в конечные продукты (при этом не происходит их включения во вновь образующиеся молекулы нуклеиновых кислот в тканях организма). То есть, нуклеиновые кислоты пищи практически не выступают в роли поставщика непосредственных предшественников нуклеиновых кислот тканей организма.
В то же время нуклеотиды и нуклеозиды, введённые парентерально, могут включаться в нуклеиновые кислоты без всяких изменений. Это послужило основой методов исследования метаболизма нуклеиновых кислот путём введения меченных радиоактивными изотопами азотистых оснований.
Распад пуриновых и пиримидиновых нуклеотидов до конечных продуктов в тканях
Распад пуриновых нуклеотидов.
Аденозин и гуанозин, которые образуются при гидролизе пуриновых нуклеотидов, подвергаются ферментативному распаду с образованием конечного продукта – мочевой кислоты, которая выводится с мочой из организма.

Распад пиримидиновых нуклеотидов.
Начальные этапы этого процесса катализируются специфическими ферментами. Конечные продукты: СО2, NН3, мочевина, β-аланин, β-аминоизомасляная кислота. β-аланин используется для синтеза дипептидов мышц – карнозина и ансерина или выделяется с мочой.
21. Коферменты: Большинство ферментов для проявления ферментативной активности нуждается в низкомолекулярных органических соединениях небелковой природы (коферментах) и/или в ионах металлов (кофакторах). Термин. «кофермент» был введён в начале XX века и обозначал часть некоторых ферментов, которая легко отделялась от белковой молекулы фермента и удалялась через полупроницаемую мембрану при диализе. Несколько позже было выяснено, что большинство ферментов состоит из термолабильной белковой части и термостабильного небелкового фактора — кофермента. Белковая часть получила название «апофермент», который в отсутствие кофермента не обладает каталитической активностью. Кофермент с белковой молекулой (апоферментом) формируют молекулу холофермента, обладающую каталитической активностью.
Более 25% всех ферментов для проявления полной каталитической активности нуждается в ионах металлов. Рассмотрим роль кофакторов в ферментативном катализе.
1. Роль металлов в присоединении субстрата
в активном центре фермента
Ионы металла выполняют функцию стабилизаторов молекулы субстрата, активного центра фермента и конформации белковой молекулы фермента, а именно третичной и четвертичной структур.
Ионы металлов — стабилизаторы молекулы субстрата
Для некоторых ферментов субстратом служит комплекс превращаемого вещества с ионом металла. Например, для большинства киназ в качестве одного из субстратов выступает не молекула АТФ, а комплекс Mg2+-ATФ. В этом случае ион Mg2+ не взаимодействует непосредственно с ферментом, а участвует в стабилизации молекулы АТФ и нейтрализации отрицательного заряда субстрата, что облегчает его присоединение к активному центру фермента

Схематично роль кофактора при взаимодействии фермента и субстрата можно представить как комплекс E-S-Me, где Е — фермент, S — субстрат, Me — ион металла.
В качестве примера можно привести расположение субстратов в активном центре гексокиназы
Гексокиназа катализирует перенос концевого, γ-фосфатного остатка молекулы АТФ на глюкозу с образованием глюкозо-6-фосфата:

Участие ионов магния в присоединении субстрата в активном центре гексокиназы. В активном центре гексокиназы есть участки связывания для молекулы глюкозы и комплекса Мд2+-АТФ. В результате ферментативной реакции происходит перенос концевого, γ-фосфатного остатка молекулы АТФ на глюкозу с образованием глюкозо-6-фосфата.
Ион Mg2+ участвует в присоединении и «правильной» ориентации молекулы АТФ в активном центре фермента, ослабляя фосфоэфирную связь и облегчая перенос фосфата на глюкозу.
Ионы металла — стабилизаторы активного центра фермента
В некоторых случаях ионы металла служат «мостиком» между ферментом и субстратом. Они выполняют функцию стабилизаторов активного центра, облегчая присоединение к нему субстрата и протекание химической реакции. В ряде случаев ион металла может способствовать присоединению кофермента. Перечисленные выше функции выполняют такие металлы, как Mg2+, Mn2+, Zn2+, Co2+, Мо2+. В отсутствие металла эти ферменты активностью не обладают. Такие ферменты получили название «металлоэнзимы». Схематично данный процесс взаимодействия фермента, субстрата и металла можно представить следующим образом:
К металлоэнзимам относят, например, фермент пируват киназу (рис. 2-4), катализирующий реакцию:

Пищеварительные ферменты:Ферме́нты пищеваре́ния, пищеварительные ферменты — ферменты, расщепляющие сложные компоненты пищи до более простых веществ, которые затем всасываются в организм. В более широком смысле пищеварительными ферментами также называют все ферменты, расщепляющие крупные (обычно полимерные) молекулы на мономеры или более мелкие части. Пищеварительные ферменты находятся в пищеварительной системе человека и животных. Кроме этого, к таким ферментам можно отнести внутриклеточные ферменты лизосом. Основные места действия пищеварительных ферментов в организме человека и животных — это ротовая полость, желудок, тонкая кишка. Эти ферменты вырабатываются такими железами, как слюнные железы, железы желудка, поджелудочная железа и железы тонкой кишки. Часть ферментативных функций выполняется облигатной кишечной микрофлорой. По субстратной специфичности пищеварительные ферменты делятся на несколько основных групп:
Читайте также: Smile dusty blue ткань
протеазы (пептидазы) расщепляют белки до коротких пептидов или аминокислот
липазы расщепляют липиды до жирных кислот и глицерина
карбогидразы гидролизуют углеводы, такие как крахмал или сахара, до простых сахаров, таких как глюкоза
нуклеазы расщепляют нуклеиновые кислоты до нуклеотидов.
1.Ротовая полость — Слюнные железы секретируют в полость рта альфа-амилазу (птиалин), которая расщепляет высокомолекулярный крахмал до более коротких фрагментов и до отдельных растворимых сахаров (декстрины, мальтоза, мальтриоза).

Ферменты, секретирующиеся желудком называются желудочными ферментами.
Пепсин — основной желудочный фермент. Расщепляет белки до пептидов.
Желатиназа расщепляет желатин и коллаген, основные протеогликаны мяса.
-Ферменты поджелудочной железы
Поджелудочная железа является основной железой в системе пищеварения. Она секретирует ферменты в просвет двенадцатиперстной кишки.
Трипсин является протеазой, аналогичной пепсину желудка.
Химотрипсин — также протеаза, расщепляющая белки пищи.
Несколько различных эластаз, расщепляющих эластин и некоторые другие белки.
Нуклеазы, расщепляющие нуклеиновые кислоты ДНК и РНК.
Стеапсин, расщепляющий жиры.
Амилазу, расщепляющую крахмал и гликоген, а также другие углеводы.
Липаза поджелудочной железы является важнейшим ферментом в переваривании жиров. Она действует на жиры (триглицериды), предварительно эмульгированные желчью, секретируемой в просвет кишечника печенью.
Несколько пептидаз, в том числе:
энтеропептидаза — превращает трипсиноген в трипсин;
аланинаминопептидаза — расщепляет пептиды, образовавшиеся из белков после действия протеаз желудка и поджелудочной железы.
Ферменты, расщепляющие дисахариды до моносахаридов:
сахараза расщепляет сахарозу до глюкозы и фруктозы;
мальтаза расщепляет мальтозу до глюкозы;
изомальтаза расщепляет мальтозу и изомальтозу до глюкозы;
лактаза расщепляет лактозу до глюкозы и галактозы.
Липаза кишечника расщепляет жирные кислоты.
Эрепсин, фермент, расщепляющий белки.
22. Полиферментная сиситема:Каждая клетка организма имеет свой специфический набор ферментов. Некоторые из них содержатся во всех клетках, другие присутствуют только в некоторых. В клетке работа каждого фермента, как правило, не индивидуальная, а тесно связана с другими ферментами, т.е. из отдельных ферментов формируются полиферментные системы, или конвейеры. Субстрат иногда во время своего превращения проходит длинную цепь реакций, в которых участвует много ферментов. Продукт реакции, катализируемой первый фермент, служит субстратом для второго фермента и т.д. Примером может служить процесс гликолиза. Все ферменты гликолиза имеющиеся в растворимом состоянии. В процессах превращения глюкозы до молочной кислоты участвует целый ряд ферментов. Положение каждого фермента в цепи устанавливается по родством с субстратами (начиная с глюкозы), каждый из которых соответственно являются продуктом реакции, катализованои предыдущим ферментом. Это увеличивает скорость ферментативных реакций, и в такой цепи промежуточные продукты не накапливаются.
Многие полиферментные ансамблей структурно связаны с какой-либо органеллы (митохондрии, рибосомы, ядро) или биомембран-ми и составляют высокоорганизованные системы, обеспечивающие жизне-воважливи функции, например, тканевое дыхание, т.е. перенос электронов и протонов от субстратов к кислорода через систему дыхательных ферментов, закрепленных на внутренней мембране митохондрий. Некоторые ферменты, участвующие в реакции одной цепи метаболизма, объединяются в мультиэнзимных комплексы с определенной функцией. Типичным примером подобных надмолекулярных комплексов является пируватдегидрогеназный комплекс, состоящий из нескольких ферментов, участвующих в окислении пировиноградной кислоты до ацетил-КоА, или синтетаза жирных кислот, состоящий из семи структурно связанных ферментов, которые выполняют функцию синтеза жирных кислот.
23. Пищеварение в метоболизме:Метаболи́зм (от греч. μεταβολή — «превращение, изменение»), или обмен веществ — набор химических реакций, которые возникают в живом организме для поддержания жизни. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды.
Пищеварение: Такие макромолекулы, как крахмал, целлюлоза или белки, должны расщепляться до более мелких единиц прежде, чем они могут быть использованы клетками. Несколько классов ферментов принимают участие в деградации: протеазы, которые расщепляют белки до пептидов и аминокислот, гликозидазы, которые расщепляют полисахариды до олиго- и моносахаридов.
Микроорганизмы выделяют гидролитические ферменты в пространство вокруг себя,[29][30] чем отличаются от животных, которые выделяют такие ферменты только из специализированных железистых клеток.[31] Аминокислоты и моносахариды, образующиеся в результате активности внеклеточных ферментов, затем поступают в клетки с помощью активного транспорта
Пищеварительные ферменты:Ферме́нты пищеваре́ния, пищеварительные ферменты — ферменты, расщепляющие сложные компоненты пищи до более простых веществ, которые затем всасываются в организм. В более широком смысле пищеварительными ферментами также называют все ферменты, расщепляющие крупные (обычно полимерные) молекулы на мономеры или более мелкие части. Пищеварительные ферменты находятся в пищеварительной системе человека и животных. Кроме этого, к таким ферментам можно отнести внутриклеточные ферменты лизосом. Основные места действия пищеварительных ферментов в организме человека и животных — это ротовая полость, желудок, тонкая кишка. Эти ферменты вырабатываются такими железами, как слюнные железы, железы желудка, поджелудочная железа и железы тонкой кишки. Часть ферментативных функций выполняется облигатной кишечной микрофлорой. По субстратной специфичности пищеварительные ферменты делятся на несколько основных групп:
протеазы (пептидазы) расщепляют белки до коротких пептидов или аминокислот
липазы расщепляют липиды до жирных кислот и глицерина
карбогидразы гидролизуют углеводы, такие как крахмал или сахара, до простых сахаров, таких как глюкоза
нуклеазы расщепляют нуклеиновые кислоты до нуклеотидов.
1.Ротовая полость — Слюнные железы секретируют в полость рта альфа-амилазу (птиалин), которая расщепляет высокомолекулярный крахмал до более коротких фрагментов и до отдельных растворимых сахаров (декстрины, мальтоза, мальтриоза).
Ферменты, секретирующиеся желудком называются желудочными ферментами.
Пепсин — основной желудочный фермент. Расщепляет белки до пептидов.
Желатиназа расщепляет желатин и коллаген, основные протеогликаны мяса.
-Ферменты поджелудочной железы
Поджелудочная железа является основной железой в системе пищеварения. Она секретирует ферменты в просвет двенадцатиперстной кишки.
Трипсин является протеазой, аналогичной пепсину желудка.
Химотрипсин — также протеаза, расщепляющая белки пищи.
Несколько различных эластаз, расщепляющих эластин и некоторые другие белки.
Нуклеазы, расщепляющие нуклеиновые кислоты ДНК и РНК.
Стеапсин, расщепляющий жиры.
Амилазу, расщепляющую крахмал и гликоген, а также другие углеводы.
Липаза поджелудочной железы является важнейшим ферментом в переваривании жиров. Она действует на жиры (триглицериды), предварительно эмульгированные желчью, секретируемой в просвет кишечника печенью.
Несколько пептидаз, в том числе:
энтеропептидаза — превращает трипсиноген в трипсин;
аланинаминопептидаза — расщепляет пептиды, образовавшиеся из белков после действия протеаз желудка и поджелудочной железы.
Ферменты, расщепляющие дисахариды до моносахаридов:
сахараза расщепляет сахарозу до глюкозы и фруктозы;
мальтаза расщепляет мальтозу до глюкозы;
изомальтаза расщепляет мальтозу и изомальтозу до глюкозы;
лактаза расщепляет лактозу до глюкозы и галактозы.
Липаза кишечника расщепляет жирные кислоты.
Эрепсин, фермент, расщепляющий белки.
24. Тканевое дыхание.Клеточное или тканевое дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды. Высвобожденная энергия запасается в химических связях макроэргических соединений (АТФ и др.) и может быть использована по мере необходимости. Входит в группу процессов катаболизма. О физиологических процессах транспортировки к клеткам многоклеточных организмов кислорода и удалению от них углекислого газа см. статью Дыхание.
Читайте также: Разъедает ли вазелин ткань
Впервые сущность дыхания объяснил А.-Л. Лавуазье (1743-1794), обративший внимание на сходство между горением органических веществ вне организма и дыханием животных. Постепенно становились ясными принципиальные различия между этими двумя процессами: в организме окисление протекает при относительно низкой температуре в присутствии воды, и его скорость регулируется обменом веществ. В настоящее время биологическое окисление определяется как совокупность реакций окисления субстратов в живых клетках, основная функция которых — энергетическое обеспечение метаболизма. В развитие концепций биологического окисления в XX в. важнейший вклад внесли А.Н. Бах, О. Варбург, Г. Крепс, В.А. Энгель-гардт, В.И. Палладин, В.А. Белицер, С.Е. Северин, В.П. Скулачев.
БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ — совокупность ферментативных окислительно-восстановительных реакций, протекающих в живых клетках. В процессе биологического окисления происходит расщепление питательных веществ, и освобождаемая при этом энергия запасается в удобной для использования клетками форме т. н. богатых энергией соединений — аденозинтрифосфатов и др. Эти соединения затем расходуются на обеспечение всех процессов жизнедеятельности; часть энергии рассеивается в виде тепла. Значительная часть реакций биологического окисления осуществляется в митохондриях
Анаэробное окисление аммония, anammox — биохимический процесс окисления иона аммония нитрит-анионом в анаэробных условиях. Служит источником энергии для фиксации углекислого газа. Описан у следующих родов бактерий: Brocadia, Kuenenia, Anammoxoglobus, Jettenia, Scalindua. Все они относятся к планктомицетам.
Процесс был открыт в 1986 году. Сейчас создана новая технология очистки сточных вод от соединений азота с помощью осуществляющих анаэробное окисление аммония бактерий. В Роттердаме (Нидерланды) построена и запущена первая очистная станция на её основе. Важными достоинствами данной технологии являются уменьшение выбросов CO2 в атмосферу на 85-90% по сравнению с традиционными методами, а также относительная дешевизна.
Общее уравнение реакций анаэробного окисления аммония:
Анаэробное окисление метана — процесс окисления метана до углекислого газа, производимый некультивируемыми (англ. VBNC) археями групп ANME-1, ANME-2 и ANME-3, близкими к Methanosarcinales, в ассоциации с сульфатредуцирующими и денитрифицирующими бактериями при отсутствии в среде молекулярного кислорода. Биохимия и распространённость процесса в природе изучены пока недостаточно.
26. Пируват, образовавшийся в реакциях гликолиза (в цитоплазме), должен быть транспортирован в митохондрии. Транспорт осуществляется специальной «челночной» системой. В матриксе митохондрии, прикрепившись к ее внутренней мембране, находится сложный полиферментный комплекс – пируватдегидрогеназа.
Пируватдегидрогеназа состоит из 60 полипептидных цепей, которые можно разделить на 3 основных фермента: Е1 – собственно пируватдегидрогеназа (состоит из 24 субъединиц); Е2 – дигидролипоилтрансацетилаза (также 24 субъединицы); Е3 – дигидролипоилдегидрогеназа (12 субъединиц).
Последовательность реакций представлена на рис.5.12. Е1 катализирует декарбоксилирование ПВК с участием кофермента тиаминпирофосфата (ТПФ). Образовавшийся продукт реакции (гидроксиэтильное производное ТПФ) при участии Е2 реагирует с окисленной липоевой кислотой(ЛК). Липоевая кислота – низкомолекулярное азотсодержащее соединение – является коферментом Е2.
Дисульфидная группа ЛК способна восстанавливаться и ацетилироваться. В реакции, катализируемой дигидролипоилтрансацетилазой (Е2 ), образуется ацетиллипоевая кислота. Далее это соединение реагирует с коэнзимом А (КоА-SH не является собственным коферментом Е2) – при этом образуется восстановленная форма ЛК (дигидролипоевая кислота) и ацетил-КоА.
Наконец, начинает функционировать Е3 , коферментом которого является ФАД: кофермент окисляет дигидролипоевую кислоту и сам при этом восстанавливается (ФАДН2) . Восстановленный флавиновый кофермент реагирует с митохондриальным НАД+, в свою очередь, восстанавливая его (НАДН ·Н+).
Таким образом, в окислительном декарбоксилировании ПВК участвует фактически три фермента, составляющих единый пируватдегидрогеназный комплекс, и 5 коферментов: ТПФ, ЛК и ФАД – собственные коферменты комплекса, КоА-SH и НАД+ – внешние, приходящие “извне”. Образующийся ацетил-КоА затем окисляется в цикле Кребса, а водород с НАДН ·Н+ поступает в дыхательную цепь митохондрий.

Механизм функционирования пируватдегидрогеназного комплекса
Пируватдегидрогеназа отличается большим отрицательным редокс-потенциалом, который способен обеспечить не только восстановление НАД+, но и способствовать образованию высокоэнергетической тиоэфирной связи в ацетил-КоА (СН3-СО
При недостаточном содержании в диете входящих в состав пируватдегидрогеназы витаминов, в первую очередь тиамина, активность фермента снижается. Это приводит к накоплению в крови и тканях пирувата и лактата и развитию метаболического ацидоза. При выраженном дефиците тиамина развивается некомпенсированный ацидоз, который без лечения приводит к летальному исходу.
^ Регуляция активности пируватдегидрогеназы
Пируватдегидрогеназный комплекс может существовать в активной и неактивной формах. Переход одной формы в другую осуществляется путем обратимого фосфорилирования с участием киназы и дефосфорилирования с участием фосфатазы. При этом фосфорилированная форма является неактивной, а дефосфорилированная – активной .
При низкой концентрации инсулина и высоком уровне энергообеспеченности клетки (↑АТФ, ↑ ацетил-КоА и ↑ НАДН·Н+) этот комплекс находится в неактивном состоянии. Активирование пируватдегидрогеназного комплекса индуцируется инсулином, КоА-SН, пируватом, АДФ и ионами магния.
28.Тканевое дыхание и биологическое окисление. Распад органических соединений в живых тканях, сопровождающийся потреблением молекулярного кислорода и приводящий к выделению углекислого газа и воды и образованию биологических видов энергии, называется тканевым дыханием. Тканевое дыхание представляют как конечный этап пути превращений моносахаров (в основном глюкозы) до указанных конечных продуктов, в который на разных стадиях включаются другие сахара и их производные, а также промежуточные продукты распада липидов (жирные кислоты), белков (аминокислоты) и нуклеиновых оснований. Итоговая реакция тканевого дыхания будет выглядеть следующим образом:
С6Н12О6 + 6O2 = 6СO2+ 6Н2O + 2780 кДж/моль. (1)
Потребление кислорода тканями зависит от интенсивности реакций тканевого дыхания. Наибольшей скоростью тканевого дыхания характеризуются почки, мозг, печень, наименьшей – кожа, мышечная ткань (в покое). Уравнение (2) описывает суммарный результат многоступенчатого процесса, приводящего к образованию молочной кислоты (см. главу 10) и протекающего без участия кислорода:
С6Н12Об = 2С3Н6О3 + 65 кДж/моль. (2)
Этот путь отражает, по-видимому, энергетическое обеспечение простейших форм жизни, функционировавших в бескислородных условиях. Современные анаэробные микроорганизмы (осуществляющие молочнокислое, спиртовое и уксуснокислое брожение) получают для жизнедеятельности энергию, производимую в процессе гликолиза или его модификаций.
Использование клетками кислорода открывает возможности для более полного окисления субстратов. В аэробных условиях продукты бескислородного окисления становятся субстратами цикла трикарбоновых кислот (см. главу 10), в ходе которого образуются восстановленные дыхательные переносчики НАДФН, НАДН и флавиновые коферменты. Способность НАД+ и НАДФ+ играть роль промежуточного переносчика водорода связана с наличием в их структуре амида никотиновой кислоты. При взаимодействии этих кофакторов с атомами водорода имеет место обратимое гидрирование (присоединение атомов водорода):
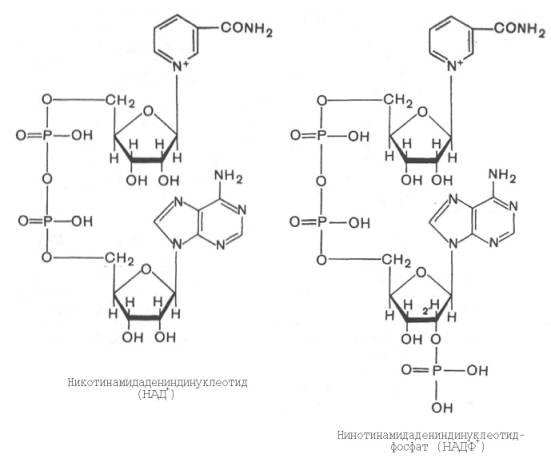
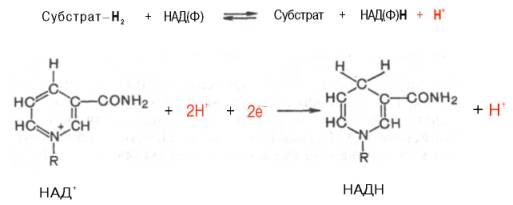
При этом в молекулу НАД+ (НАДФ+) включаются 2 электрона и один протон, а второй протон остается в среде.
Во флавиновых коферментах (ФАД или ФМН), активной частью молекул которых является изоаллоксазиновое кольцо, в результате восстановления чаще всего наблюдается присоединение 2 протонов и 2 электронов одновременно:


Восстановленные формы этих кофакторов способны транспортировать водород и электроны к дыхательной цепи митохондрий или иных энерго-сопрягающих мембран