
Жирные кислоты входят в состав большей части липидов. Поэтому вопросы обмена липидов – это, как правило, вопросы обмена жирных кислот: их источники и пути дальнейших превращений.
Судьба жирной кислоты зависит от ее строения(насыщенная ЖК или полиненасыщенная ЖК) и от внутриклеточных условий(наличие или отсутствие энергии).
Состояние покоя и отдыха в абсорбтивный период
В течение нескольких часов после приема пищи (абсорбтивный период) экзогенныенасыщенныеимононенасыщенныежирные кислоты поступают с помощьюхиломикронов из кишечника в ткани, которые имеют липопротеинлипазуна эндотелии капилляров.
Параллельно в этот период времени в печени жирные кислоты способны синтезироваться из избытка экзогенной глюкозы и полученные эндогенные жирные кислоты этерифицируются с глицеролом в реакцияхлипогенезас образованием ТАГ. Далее они транспортируются из печени в ткани, имеющие липопротеинлипазу, в составе ЛПОНП.
В клетках жировой ткани после приема пищи насыщенная жирная кислота либо синтезируется из глюкозы, либо поступает из хиломикронов и ЛПОНП. Далее она направляется в липогенез и запасается в составе ТАГ.
Еслилипопротеинлипазы нетв ткани, то жирная кислота доставляется сюда в комплексе с альбумином(как в случае со скелетной мышцей). Этот комплекс образуется после работы липопротеинлипазы в других тканях.
Полиненасыщенныежирные кислоты (ПНЖК) обычно поступают в клетки в виде эфиров холестерола ифосфолипидовв составе ЛПВП и ЛПНП. Эти жирные кислоты необходимы для синтеза эйкозаноидовв некоторых видах клеток либо они участвуют в синтезе фосфолипидов для мембран клетки.

Возможные источники и пути использования жирной кислоты в клетке
Голодание, мышечная работа, покой в постабсорбтивный период
В постабсорбтивный период и при голоданиихиломикроны и ЛПОНП в крови отсутствуют. Так как данному состоянию обычно сопутствует гипогликемия, то для ее компенсации из поджелудочной железы секретируется глюкагон. Под влиянием глюкагона и других гормонов в жировых депо активируется расщепление ТАГ до жирных кислот и глицерола (липолиз). Транспорт жирных кислот, вышедших в кровь, осуществляется альбумином.
При физической работе гормоны адреналин, соматотропин и глюкокортикоиды, влияя на адипоциты, также вызывают в них липолизи выход жирных кислот в кровь. В комплексе с альбумином эти кислоты главным образом доставляются в мышцу для обеспечения мышечного сокращения.
В состоянии покоя, когда процессы пищеварения уже закончились, при краткосрочноми длительномголодании, прифизической нагрузке в большинстве клеток, кроме нейронов и эритроцитов, жирные кислоты сгорают в процессах β-окисленияи ЦТК, обеспечивая 50% и более всей энергии клетки.
В печени при длительном голодании (более 20 часов) жирные кислоты направляются в кетогенезна синтез кетоновых тел. Кетоновые тела далее разносятся по организму, преобразуются в ацетил-SКоА и используются для энергетического обеспечения клеток.
Триацилглицеролы нужны при нагрузке
Синтезируясь во время и сразу после приема пищи (липогенез) и запасаясь в жировой ткани, триацилглицеролы являются формой хранения насыщенных и мононенасыщенных жирных кислот. Распад триацилглицеролов (триглицеридов) по-другому называется липолиз или мобилизация жира. Он идет в жировых клетках постоянно и обычно существует равновесие между синтезом и распадом ТАГ.
Даже в состоянии покоя организма печень, сердце, скелетные мышцы и другие ткани (кроме эритроцитов и нейроцитов) более 50% энергии получают из окисления жирных кислот, поступающих из жировой ткани благодаря фоновому липолизу. По мере уменьшения резервов глюкозы клетки все больше энергии получают из окисления жирных кислот. Таким образом, насыщенные жирные кислоты выполняют роль своеобразного буфера энергии в организме.
Мобилизация триацилглицеролов и окисление жирных кислот активируется
- при нормальных физиологических стрессовых ситуациях – эмоциональный стресс, мышечная работа,голодание,
- при патологических состояниях – сахарный диабет I типа, другие гормональные заболевания (гиперкортицизм, гипертиреоз).
В результате липолизав адипоцитах образуются свободный глицероли жирные кислоты.
Глицеролс кровью доставляется в печень и почки, здесь фосфорилируется и окисляется в метаболит гликолиза диоксиацетонфосфат. В зависимости от условий ДАФ может включаться в реакцииглюконеогенеза(при голодании, мышечной нагрузке) или окисляться в гликолизе до пировиноградной кислоты.
Жирные кислоты транспортируются в крови в комплексе с альбуминами плазмы:
- при физической нагрузке – в мышцы,
- в обычных условиях и при голодании – в мышцы и большинство тканей, однако при этом около 30% жирных кислот захватывается печенью.
При голоданиии физической нагрузке после проникновения в клетки жирные кислоты вступают на путь β-окисления.
В целом мобилизацию жира можно представить как последовательность следующих событий:
- Липолиз– гормонзависимый распад ТАГ в жировой ткани или резервных ТАГ в самой клетке.
- Транспортжирных кислот из жировой ткани по крови в комплексе с альбумином.
- Проникновение жирной кислоты в цитозоль клетки-мишени.
- Активацияжирной кислоты через присоединение HS-КоА.
- Карнитин-зависимое перемещение жирной кислоты в митохондрию.
- Окислениежирной кислоты с образованием ацетильных групп (в форме ацетил-S-КоА).
- Сгорание ацетил-S-КоА в цикле лимонной кислоты или синтез (только в печени) кетоновых тел.

Обмен таг. Распад таг.
ТАГ поэтапно расщепляется соответственно тканевым липидами. Липазы делят по продуктам распада, который синтезируется.
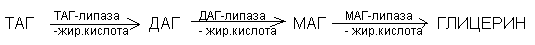
Ключевым ферментом является гормонально зависимая ТАГ-липаза. Образующийся на этом этапе глицерин и жирные кислоты оказываются в тканях с целью образования Е.
Разделяют несколько вариантов окисления жирных кислот в зависимости от углеродного атома: α-окисление, β-окисление, ω-окисление. Основным вариантом является β-окисление. Наимболее активно оно протекает в жировой ткани, печени, почках и сердечной мышце.
β-окисление заключается в постепенном отщеплении от жирной кислоты двух углеродных фрагментов в виде ацетил КоА с освобождением энергии. Запас жирных кислот расположенных в цитозоле, в котром протекает и активация жирных кислот с образованием из них ацетил КоА.
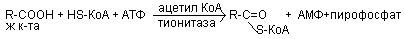
Последующее β-окисление ацетил КоА идет в митохондриях. Митохондриальная мембрана не проницаема для длинноцепочных ацетил КоА. В переносе их внутрь митохондрий участвует специальный переносчик карнитин (метил, гидропроизводной аминомасляной кислоты). Суть переноса: ацетил КоА взаимодействует с карнитином образует комплекс.

Химизм митохондриального β-окисления.

Энергитическая эффективность- бета окисления складывается из:
а) Е окисления ацетил КоА в цикле Кребса
б) Е самого бета окисления
Энергия оскисления тем выше, чем длинее углеродная цепь. Количество молекул ацетил КоА из данной жирной кислоты определяется по формуле:
1) n=Nc/2, где n-количество атомов, N- число С атомов кислоты.
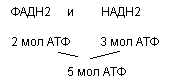
Итого: 17(N/2)-7 – бета окисление насыщенной кислоты с четным числом атомов «С». При окислении жирных кислот с нечетным числом «С» образуется сукцинил КоА который вступает в цикл Кребса.
Особенности окисления ненасыщенных жирных кислот
Начальные стадии окисления – обычное бета окисление до места двойной связи. Затем если эта двойная связь находится в альфа или бета положении, то продолжается окисление жирной кислоты со 2 этапа (минуя стадию восстановления ФАД→ФАДН2. Но если двойная связь находится не в альфа или бета положении, то ферментами енолтрансферазами связь перемещается в альфа или бета положение т.о. при окислении ненасыщенных жирных кислот образуется меньше энергии по формуле: 7(N/2)-7-2m, где m-число двойных связей, теряется образование ФАДН2.
На первом этапе глицерин активируется:
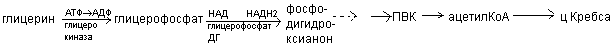
Для синтеза ТАГ нужен глицерин. Если глицерин не поставряется с пищей, то может синтезироваться из углеводов по обратным реакциям.
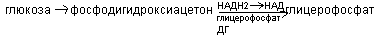
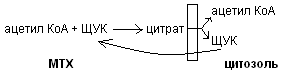
1. Синтез насыщенных жирнык кислот в цитозоле при участии сложного полиферментного комплекса (синтетаза жирных кислот или пальматат синтетаза). Этот комплекс включает особый ацил-переносящий белок и 6 ферментов. Для синтеза жирных кислот донором Н является НПДФН2, образовавшийся в пентозо-фосфатном пути. Источником для синтеза жирных кислот является ацетил КоА, который образуется в митохондриях и самостоятельно выйти в цитозоль не может. Транспорт ацетил КоА из митохондрий в цитозоль осуществляется с помощю цитратно-челночного механизма:
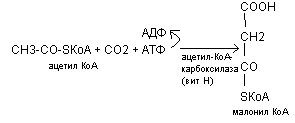
Ацетил КоА на 1 этапе карбоксилируется (+СО2) в молонил КоА.
Ацетил КоА и малонил КоА содержат полиферментный комплекс (синтетаза жирных кислот), в котором имеется 2SН-группы.


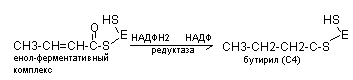
В последующем к этому комплексу присоединяется новая молекула малонил КоА и в результате синтезируется капроновая кислота (+2 «С») и далее до С16 (пальмитиновая кислота) с участием элонгазы.
2. Синтез ненасыщенных жирных кислот. У человека синтезируется только мононенасыщенные кислоты. Они оборазуются из соответствующих насыщенных жирных кислот в лизосомах при участии десатураз (ферментативной моносинтетазы, которые требуют цитохромов Р450).
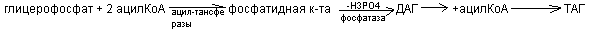
Синтезируется из фосфатидной кислоты и азотсодержащих добавочных веществ. Активатором в синтезе фосфолипидов служит ЦТФ, который может активировать фосфатидную кислоту или серин.
а) Активация фосфатидной кислоты:
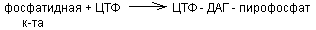

б) активация холина, серина, коламина:
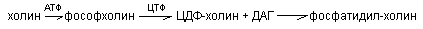
т.о. синтез фосфолипидов происходит вначале одинаково, т.е. через стадию фосфатидной к-ты. Для усиления активации синтеза структурных фосфолипидов используются липотропные вещества. К ним относится холин, метионин, вит В12 и др. Их полезно назначить после болезни Боткина для предотвращения дистрофии печени.
Осуществляется тканевыми фосфолипидами. Различают несколько видов фосфолипидов, которые разрывают различные связи.
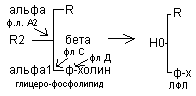
В основе ряда патологических состояний лежит тканевая активация флА2. Тогда при неполном расщеплении фосфолипида образуется лизофосфолипид.
а) образуются в кишечнике где они являются ПАВ и способствуют эмульгированию жиров
б) образуется в кровяном русле при обмене липопротеидов под действием ЛХАТ (лецитин-холистерол-ацил-трансфераза).

в) образуются в тканях при тканевом распаде .
Лизофосфолипиды отмечаются высокой гидрофильностью по сравнению с ФЛ. Появление лизофосфолипидов повышает проницаемость клеточной мембраны для Na + , К + , Н2О в результате клетки подвергаются разбуханию и разрыву. В эритроцитах наблюдается гемолиз. Фосфолипаза А2 отсутствует в яде ряда змей.
Распад таг в тканях
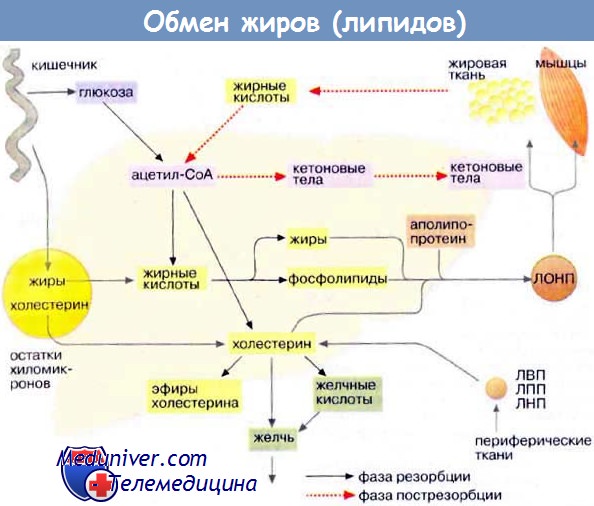
После всасывания в эпителий кишки свободные жирные кислоты и 2-моноглицериды вновь образуют триглицериды и вместе с фосфолипидами и холестеролом включаются в хиломикроны. Хиломикроны транспортируются с током лимфы через грудной проток в верхнюю полую вену, попадая таким образом в общий кровоток.
Внутри хиломикрона триглицериды гидролизуются липопротеинлипазой, что приводит к высвобождению жирных кислот на поверхности кровеносных капилляров в тканях. Это обусловливает транспорт жирных кислот в ткани и последующее образование остатков хиломикронов, обедненных триглицеридами. Эти остатки затем забирают эфиры холестерола из липопротеинов высокой плотности, и частицы быстро захватываются печенью. Данную систему транспорта жирных кислот пищевого происхождения называют экзогенной транспортной системой.
Также существует эндогенная транспортная система, предназначенная для внутриорганного транспорта жирных кислот, образованных в самом организме. Липиды транспортируются из печени в периферические ткани и обратно, а также переносятся из жировых депо к различным органам. Транспорт липидов от печени к периферическим тканям включает согласованные действия ЛПОНП, липопротеинов промежуточной плотности (ЛППП), липопротеинов низкой плотности (ЛПНП) и липопротеинов высокой плотности (ЛПВП). Частицы ЛПОНП подобно хиломикронам состоят из большого гидрофобного ядра, образованного триглицеридами и эфирами холестерола, и поверхностного липидного слоя, состоящего в основном из фосфолипидов и холестерола.
ЛПОНП синтезируются в печени, и отложение жира в периферических тканях является их основной функцией. После попадания в кровоток ЛПОНП подвергаются воздействию липопротеинлипазы, которая гидролизует триглицериды до свободных жирных кислот. Свободные жирные кислоты, происходящие из хиломикронов или ЛПОНП, могут быть использованы в качестве источников энергии, структурных компонентов фосфолипидных мембран или превращаться обратно в триглицериды и в таком виде сохраняться. Триглицериды хиломикронов и ЛПОНП также подвергаются гидролизу липазы печени.
Частицы ЛПОНП посредством гидролиза триглицеридов превращаются в более плотные, меньшие по размеру холестерол- и триглицерид-обогащенные остатки (ЛППП), которые удаляются из плазмы с помощью рецепторов печеночных липопротеинов или могут быть превращены в ЛПНП. ЛПНП являются основными липопротеинами-переносчиками холестерола.
Возврат липидов из периферических тканей в печень часто называют обратным транспортом холестерола. Частицы ЛПВП участвуют в этом процессе, забирая холестерол из тканей и других липопротеинов и перенося его в печень для последующей экскреции. Еще один вид транспортировки, существующей между органами, — перенос жирных кислот из жировых депо к органам для окисления.
Жирные кислоты, получаемые в основном в результате гидролиза триглицеридов жировой ткани, секретируются в плазму, где соединяются с альбумином. Связанные с альбумином жирные кислоты переносятся по градиенту концентрации в ткани с активным метаболизмом, где и используются преимущественно в качестве источников энергии.
В течение последних 20 лет только несколько исследований были посвящены вопросу транспорта липидов в перинатальном периоде (результаты этих исследований в данном издании не представлены). Очевидна необходимость более детального изучения этой проблемы.
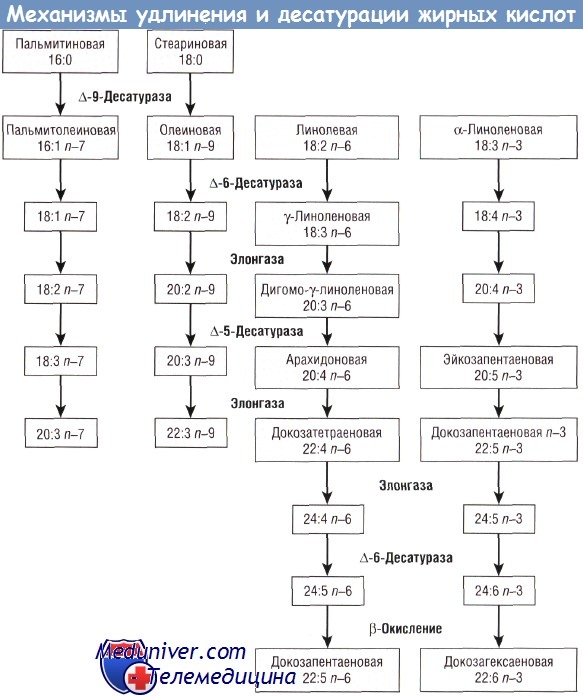
Жирные кислоты используются в качестве строительного материала в составе липидов клеточной стенки, в качестве источников энергии, а также откладываются «про запас» в виде триглицеридов преимущественно в жировой ткани. Некоторые омега-6 и омега-3 ДЦПНЖК являются предшественниками биологически активных метаболитов, используемых в сигнальной системе клеток, регуляции генов и других метаболически активных системах.
Вопрос о роли ДЦПНЖК ARA и DHA в процессе роста и развития ребенка является одним из важнейших вопросов в исследованиях, проводимых в области детской нутрициологии в течение последних двух десятилетий.
Липиды являются одними из основных компонентов клеточных мембран. Значительное количество исследований в области физиологии липидов посвящено двум жирным кислотам — ARA и DHA. ARA обнаружена в составе клеточных мембран всех структур организма человека; она является предшественником эйкозаноидов 2-й серии, лейкотриенов 3-й серии и других метаболитов, которые включены в сигнальные системы клеток и процесс генной регуляции. Исследования, посвященные DHA, часто указывают на ее структурную и функциональную роль в составе клеточных мембран.
Эта жирная кислота обнаружена в высокой концентрации в сером веществе головного мозга, а также в палочках и колбочках сетчатки. Исследования постепенного исключения из диеты животных омега-3 жирных кислот показали, что содержащие 22 атома углерода омега-6 ДЦПНЖК (например, 22:5 п—6) способны структурно, но не функционально заменить 22:6 n-3. При неадекватном уровне 22:6 n-3 в тканях выявляются нарушения зрения и познавательных способностей. Было показано, что изменение содержания 22:6 n-3 в тканях влияет на нейротрансмиттерную функцию, активность ионных каналов, сигнальные пути и экспрессию генов.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «физиология человека»