
В 1924-1928 гг. немецкие ученые Дж. Будер и А. Шмидт разработали теорию роста стебля (теорию туники и корпуса), отличающуюся от гистогенной теории Р. Ганштейна (от греч. histos — ткань и genos — род, происхождение). Согласно этой теории, в конусе нарастания стебля покрытосеменных растений выделяются две зоны: наружная — туника и внутренняя — корпус. Туника состоит из нескольких слоев клеток, чаще из двух, которые делятся перпендикулярно поверхности органа. Самый поверхностный ее слой дает начало протодерме, из которой в дальнейшем развивается эпидерма, покрывающая листья и стебли. Внутренний слой (или слой туники) образует все ткани первичной коры.
Иногда внутренние слои туники могут образовывать только наружную часть первичной коры. В таком случае происхождение внутренней части первичной коры связано с корпусом.
Это свидетельствует об отсутствии резкой границы между туникой и корпусом. Теория туники и корпуса объясняет и формирование органов побега: листьев и пазушных почек. Так, зачатки листьев закладываются во втором слое туники, а пазушные почки — в корпусе.
Развитие стебля осуществляется за счет дифференциации клеток туники и корпуса — первичных меристем. Из них образуются первичная покровная ткань — эпидерма, первичная кора и центральный осевой цилиндр (табл. 3.2).
Таблица3.2. Структура меристем стебля
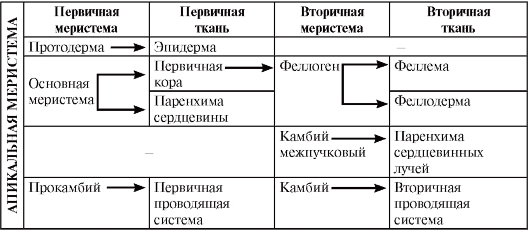
Нарастание побега осуществляется за счет апикальных меристем. Апекс побега отличается от апекса корня отсутствием чехлика и наличием листовых зачатков (примордиев). Гладким остается лишь кончик апекса, который называется конусом нарастания. В апексе побега имеются инициальные клетки, делящиеся непрерывно. У многих споровых имеется лишь одна инициаль тетраэдрической формы. У семенных растений апекс, как правило, представлен массивом инициальных клеток. В апексе цветковых растений выделяют два участка инициалей. На дистальном конце апекса хорошо выделяются два (реже 1–4) слоя клеток, делящихся антиклинально (перпендикулярно поверхности) и обеспечивающих рост апекса в поверхность. Эта зона инициальных клеток называется туникой. Несколько слоев клеток, лежащих под туникой, делящихся во всех направлениях и увеличивающих объем апекса, называют корпусом. Там, где должен возникнуть листовой зачаток, деление клеток усиливается, они становятся периклинальными; стираются границы между внутренним слоем туники и корпусом. Эта меристема называется периферической (область инициального кольца). В центре оси апекса обособляется стержневая меристема, образующая сердцевину.
Первичное строение стебля формируется за счет деятельности первичных меристем апекса. В первичном строении выделяют три анатомотопографические зоны:
3) центральный осевой цилиндр (рис. 3.27, 3.28).
Поверхность стебля однодольных растений покрыта однослойной эпидермой, которая впоследствии покрывается кутикулой. Непосредственно под эпидермой расположена первичная кора. Она представлена однородными клетками хлорофиллоносной паренхимы, граничащей со склеренхимой перициклического происхождения центрального осевого цилиндра. Перициклическая склеренхима придает растению прочность. Иногда хлорофиллоносная паренхима отсутствует, и тогда перициклическая склеренхима располагается сразу под эпидермой.
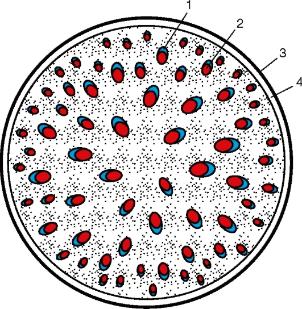
Рис. 3.27.Схема строения стебля однодольного растения: 1 — флоэма; 2 — ксилема; 3 — эпидерма; 4 — механическое кольцо
Весь центральный осевой цилиндр пронизан изолированными сосудисто-волокнистыми пучками, которые образуются за счет деятельности прокамбия. Дифференциация клеток прокамбия происходит центростремительно: флоэма формируется от периферии пучка к его центру, а ксилема — навстречу ей. У однодольных растений прокамбий полностью дифференцируется в первичные проводящие элементы (у двудольных растений прокамбиальные клетки в центре пучка формируют камбий). Форма пучков на поперечном срезе овальная: к периферии стебля расположены элементы первичной флоэмы, а к центру — первичной ксилемы. У стеблей однодольных растений формируются пучки коллатерального типа, всегда закрытые, поэтому стебель к дальнейшему утолщению не способен. Сформировавшиеся сосудисто-волокнистые пучки на срезе расположены беспорядочно в центральном осевом цилиндре. Как правило, пучки окружены склеренхимой, максимальное количество которой сосредоточено возле поверхности стебля. От периферии к центру стебля размер пучков увеличивается. Пространство между пучками занято запасающей или основной паренхимой (рис. 3.29, 3.30). Клетки основной паренхимы крупные, среди них могут быть межклетники.
Читайте также: Почему собака есть ткань
Для однодольных растений, в отличие от двудольных, нехарактерно наличие сердцевины в центре стебля, хотя может быть развита центральная воздушная полость (например, у стеблей злаков — соломина) (рис. 3.31, 3.32). Соломина — особый тип стебля с полыми междоузлиями и узлами между ними (рис. 3.33).
В зрелой соломине ржи, пшеницы и других злаков эпидерма и хлорофиллоносная паренхима, утратившие хлоропласты, подвергаются одревеснению. Это происходит к моменту созревания зерна для придания механической прочности стеблю, который приобретает в этот период вместо зеленой желтую окраску. Пучки расположены в два слоя в шахматном порядке и окружены склеренхимой. Внутренние пучки более крупные, наружные мелкие, и их склеренхимная обкладка сливается с перициклической склеренхимой, образуя кольцо механической ткани.
Для стеблей однодольных растений характерны:
• сохранение у травянистых однодольных растений первичного строения в течение всей жизни;
• слабовыраженная первичная кора;
• разбросанное расположение сосудисто-волокнистых пучков (по спирали);
• коллатеральные пучки только закрытого типа (без камбия);
• наличие во флоэме только проводящих элементов — ситовидных трубочек с клетками-спутницами;
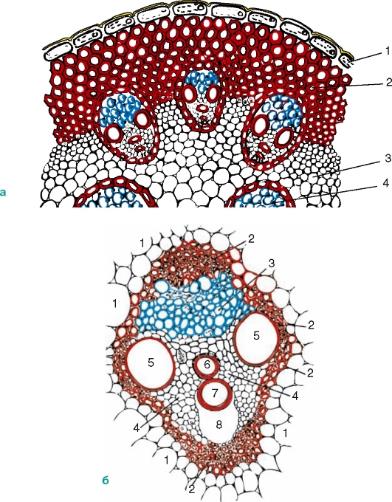
Рис. 3.28.Поперечные срезы: а— стебля кукурузы: 1 — эпидерма; 2 — склеренхима; 3 — основная паренхима; 4 — закрытый коллатеральный пучок; б— закрытого сосудисто-волокнистого пучка кукурузы: 1 — тонкостенная паренхима стебля; 2 — склеренхима; 3 — луб (флоэма); 4 — древесинная паренхима; 5 — сетчатые сосуды; 6 — кольчато-спиральный сосуд; 7 — кольчатый сосуд; 8 — воздушная полость
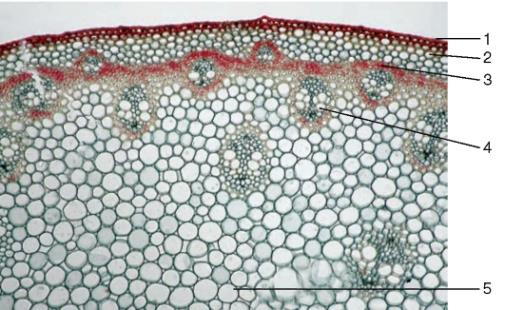
Рис. 3.29.Стебель однодольного растения (купены): 1 — одревесневшая эпидерма; 2 — паренхима первичной коры; 3 — перициклическая склеренхима; 4 — закрытый коллатеральный пучок; 5 — основная паренхима
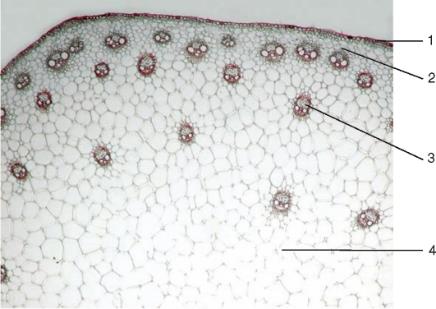
Рис. 3.30.Строение стебля однодольного растения (кукурузы): 1 — эпидерма; 2 — паренхима первичной коры; 3 — закрытый коллатеральный пучок; 4 — основная паренхима центрального осевого цилиндра
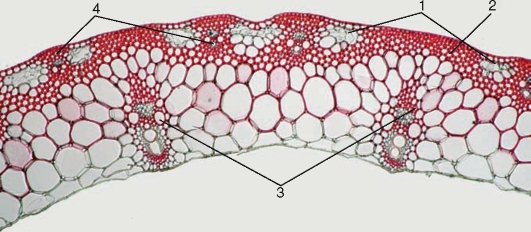
Рис. 3.31.Строение стебля соломины риса: 1 — участки хлорофиллоносной паренхимы; 2 — склеренхима; 3 — крупные пучки (мелкие и крупные пучки расположены в шахматном порядке); 4 — мелкие пучки
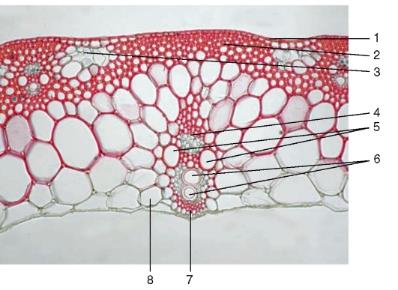
Рис. 3.32.Стебель однодольного растения соломины ржи (увеличенный фрагмент): 1 — одревесневшая эпидерма; 2 — склеренхима; 3 — хлорофиллоносная паренхима; 4 — флоэма; 5 — метаксилема; 6 — протоксилема; 7 — склеренхимная обкладка пучка; 8 — основная паренхима
Вторичное утолщение стеблей древесных однодольных растений осуществляется за счет кольца утолщения, которое представляет собой особый валик вокруг конуса нарастания, дающий дополнительный ряд сосудисто-волокнистых пучков. Подобное утолщение наблюдается у таких однодольных растений, как пальма, банановое дерево, алоэ.
Ниже рассмотрены особенности строения корневищ однодольных растений. Корневища, будучи подземным видоизменением побега, сохраняют в своем анатомическом строении характерные черты стеблей и приобретают некоторые особенности, связанные с подземным существованием. Покровной тканью остается эпидерма, часто одревесневшая. Первичная кора значительно шире и представлена запасающей паренхимой. Во внутреннем слое первичной коры, примыкающей к центральному осевому цилиндру, формируется однослойная эндодерма (подковообразная или с пятнами Каспари). Изредка, как, например, в корневище ландыша, она бывает двухслойной (рис. 3.34).
Центральный осевой цилиндр начинается с живого перицикла. Его роль в подземных побегах заключается в формировании придаточных корней. Встречаются пучки двух типов: закрытые коллатеральные иконцентрические, расположенные также беспорядочно в центральном осевом цилиндре (рис. 3.35).
Читайте также: Особенности строения человеческих тканей
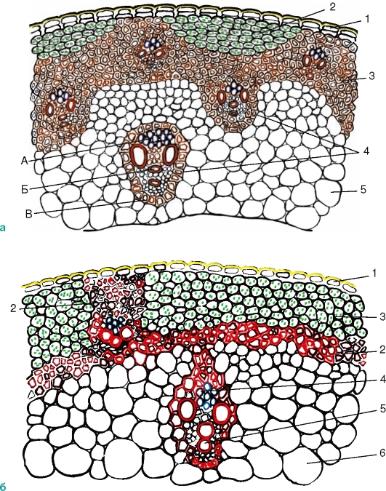
Рис. 3.33.Строение соломины: а— ржи: 1 — эпидерма; 2 — хлорофиллоносная ткань; 3 — склеренхима; 4 — закрытые коллатеральные сосудисто-волокнистые пучки: А — флоэма; Б — ксилема; В — склеренхимная обкладка пучка; 5 — основная паренхима; б— пшеницы: 1 — эпидерма; 2 — склеренхима; 3 — хлоренхима; 4 — флоэма; 5 — ксилема; 6 — основная паренхима
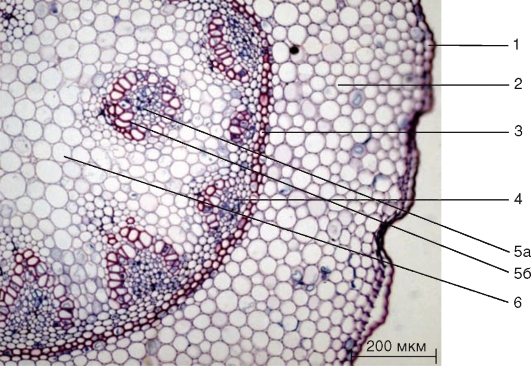
Рис. 3.34.Поперечный срез корневища ландыша: 1 — эпидерма; 2 — запасающая паренхима первичной коры; 3 — эндодерма с подковообразными утолщениями; 4 — закрытый коллатеральный пучок; 5 — концентрический пучок: 5а — флоэма; 5б — ксилема; 6 — основная паренхима
Дата добавления: 2016-01-03 ; просмотров: 3659 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Первичное строение стебля
Ниже конуса нарастания происходит активное формирование и рост постоянных тканей. У всех семенных растений стебель первичного строения состоит из 3 анатомо-топографических зон:
3) Центрального цилиндра (стели).
Первичная кора – часть стебля, между эпидермой и центральным цилиндром. Всегда живая, деятельная часть стебля. Обычно первичная кора состоит из паренхимы, но иногда в её составе может быть и механическая ткань: у двудольных – колленхима, у однодольных – склеренхима.
Первичная кора, находясь на периферии стебля, воспринимает солнечный свет и почти всегда имеет в своём составе хлоренхиму, особую разновидность паренхимы, клетки которой содержат хлоропласты и способны к фотосинтезу (поэтому молодые стебли зелёные). Она обычно расположена сразу под эпидермой. Часто роль хлоренхимы выполняет колленхима, которая дополнительно несёт механическую функцию и не препятствует росту стебля.
В средней части первичной коры обычно расположена запасающая паренхима.
Самый внутренний слой первичной коры – эндодерма состоит из крупных клеток, заполненных крахмалом, и называется крахмалоносное влагалище. В стебле этот слой не всегда хорошо выражен.
Под эндодермой находится центральный цилиндр или стела (стель).
Это центральная часть стебля (или корня) первичного строения, состоящая из проводящих тканей, паренхимы сердцевины и перицикла.
1) Наружная часть центрального цилиндра – перицикл (боковая остаточная меристема). Он быстро расходуется и в первичном стебле образует волокна склеренхимы и паренхиму или только паренхиму. У некоторых растений в стебле перицикла нет. Особенно активен перицикл у однодольных растений, он всегда формирует сплошное кольцо склеренхимы на периферии центрального цилиндра.
2) Главная часть центрального цилиндра – флоэма и ксилема, образованные за счёт работы прокамбия. Именно от их заложения, развития и расположения в стебле зависит строение центрального цилиндра.
Бывает 2 основных случая заложения флоэмы и ксилемы.
1. Прокамбий закладывается в виде отдельных продольных тяжей, в результате образуются сосудисто-волокнистые проводящие пучки, состоящие из флоэмы и ксилемы (обычно коллатеральные или биколлатеральные). Между пучками находятся радиальные полосы паренхимных клеток – сердцевинные лучи. Такой тип центральногоцилиндра называется пучковым.
Стебли двудольных и однодольных растений различаются по расположению пучков на поперечном срезе стебля:
А) у двудольных – коллатеральные или биколлатеральные открытые пучки располагаются кольцом вокруг паренхимной сердцевины или полости в центре стебля. Стелу (центр. цилиндр) такого строения называют эустелой. (Эустела может иметь и непучковое строение).
Б) у большинства однодольных – проводящие пучки располагаются диффузно по всему центральному цилиндру. Они могут быть и в перицикле, и тогда окружены склеренхимой, которую формирует перицикл, и иногда даже заходят в первичную кору. Пучки у однодольных коллатеральные или концентрические амфивазальные. ВСЕГДА ЗАКРЫТЫЕ! В стеблях однодольных растений вторичного утолщения нет, так как никогда не бывает камбия! Стелу такого строения называют атактостелой.
Читайте также: Сумка планшет из ткани своими руками
2. Непучковый (или сплошной) тип центрального цилиндра возникает тогда, когда прокамбий закладывается сплошным кольцом и формирует сплошное кольцо флоэмы и сплошное кольцо ксилемы. Сердцевинные лучи есть, но они очень узкие, незаметные, состоят из 1-2 рядов паренхимных клеток. Непучковый тип центрального цилиндра формируется только у двудольных!
3) В центре стебля находится сердцевина. У первичного стебля сердцевина хорошо развита, занимает большой объем. Она состоит из малоспециализированных паренхимных клеток, выполняющих запасающую функцию. Клетки сердцевины быстро прекращают делиться и при росте стебля в толщину часто разрываются, тогда на месте сердцевины образуется центральная полость.
Таким образом, на поперечном срезе стебля хорошо видно, что наибольший объём в нём занимает центральный цилиндр, на периферии которого находятся проводящие и механические элементы – флоэма и ксилема. Первичная кора занимает небольшой объём. Таким образом, достигается наилучшая механическая прочность и стебель хорошо противостоит нагрузкам (в основном на излом).
На продольном разрезе стебля первичного строения хорошо видно, что в местах прикрепления листьев – узлах, проводящие ткани листа проникают в стелу стебля. Эта единая проводящая система формируется уже в апексе при заложении узлов с листовыми зачатками и почками.
Из основания листа в стебель могут входить один или 3-5 (чаще нечётное число) тяжей прокамбия, которые затем дифференцируются в проводящие пучки.
Совокупность пучков, входящих из листа в стебель, до места их слияния с проводящей системой стебля называется листовым следом. Ниже из проводящих пучков листа и стебля формируются общие пучки растущего побега, входящие в его проводящую систему. Из листа в стебель приходят вещества-ассимиляты, гормоны, стимулирующие развитие стебля, то есть листовые следы оказывают большое влияние на формирование стелы стебля. У растений с пучковым типом центрального цилиндра вхождение и слияние следов происходит легко между клетками паренхимы, окружающей пучки.
У двудольных растений со сплошным типом центрального цилиндра всё гораздо сложнее.
Так как через сплошное кольцо флоэмы и ксилемы пучки свободно пройти не могут, поэтому, уже при формировании постоянных тканей, в месте вхождения следа не развиваются проводящие ткани, а вместо них образуются клетки живой паренхимы. Т.о, на поперечном и продольном срезах виден прорыв в сплошном кольце флоэмы и ксилемы. Такие прорывы называются листовыми прорывами или листовыми лакунами.
Таким же образом происходит соединение поводящей системы почек и развивающихся из них боковых ветвей. Их следы называются следами ветвления (или стеблевыми следами), а прорывы соответственно – прорывы ветвления (или стеблевые прорывы).
Ниже места вхождения следов ветвления и листовых следов стебель утолщается, за счет входящих в него дополнительных проводящих тканей.
Образование листовых и стеблевых прорывов в сплошной проводящей системе стебля сыграло огромную роль в эволюции стелы первичного стебля. Произошёл переход от сплошного центрального цилиндра к пучковому, более физиологически и энергетически выгодному.
Тот или иной путь и характер расположения листовых и стеблевых следов, число проводящих пучков в следе закреплены генетически и являются важным систематическим признаком, позволяющим судить о родстве разных групп растений.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом