
• Бластоциста — следующий после морулы этап эмбриогенеза, когда клетки морулы выделяют жидкость и формируют заполненную жидкостью полость, в которой обособляется отдельная группа клеток — внутренняя клеточная масса (эмбриобласт). Остальные клетки бластоцисты (трофобласт) формируют хорион, часть плаценты и оболочки, в которых из внутренней клеточной массы развивается эмбрион.
• Внутренняя клеточная масса (эмбриобласт) — компактная группа клеток морулы, формирующая эмбрион. Эмбриобласт расслаивается на эпибласт (слой цилиндрических клеток) и гипобласт (слой кубических клеток, обращенный к бластоцелю). Эпибласт и гипобласт вместе образуют двуслойный зародышевый диск (бластодиск).
• Гаструляция — этап развития, начинающийся после имплантации — морфогенетический процесс, в ходе которого из однослойного эпибласта образуются три первичных зародышевых листка: энтодерма, мезодерма и эктодерма; еще до начала гаструляции клетки эмбриона теряют свойство тотипотентности, т.е. способность развиваться в разных направлениях.
• Гипобласт — часть клеток внутренней клеточной массы (первичная энтодерма), формирующая оболочку плода (амнион).
• Детерминация — этап развития, на котором клетка выбирает один из многих возможных путей развития, т.е. становится предопределенной к развитию в конкретном направлении.
• Дифференцировка — приобретение клеткой новых характеристик, специфичных для конкретного типа клеток или ткани.
• Дихориальные близнецы — монозиготные близнецы, возникающие из-за разделения эмбриона на две части до стадии бластоцисты; в итоге формируются две бластоцисты.
• Зигота — оплодотворенная яйцеклетка, первый шаг в эмбриогенезе.
• Клетка-предшественница (прогениторная). Клетка, прошедшая путь развития к полностью дифференцированной клетке.
• Листки зародышевые — три четких слоя клеток, возникающие во внутренней клеточной массе: эктодерма, мезодерма и энтодерма, развивающиеся в различные ткани эмбриона.
• Мезодерма — первичный эмбриональный листок, образующий соединительную ткань, мышцы, кости, сосуды лимфатической и кроветворной системы.
• Мозаик — индивидуум, который развивается из одного оплодотворенного яйца, но в котором мутация после оплодотворения приводит к клеткам с двумя и более генотипами. Противоположность — химера.
• Мозаичное развитие — стадия развития, когда клетки коммитированы, так что удаление части эмбриона уже не приводит к нормальному развитию.
• Моноамниотические близнецы — монозиготные близнецы, появившиеся вследствие разделения части внутренней клеточной массы (эмбриобласта) без разделения части внутренней клеточной массы, формирующей амниоти-ческую оболочку (гипобласт).
• Монозиготные близнецы — близнецы, возникающие из единственной оплодотворенной яйцеклетки вследствие физического разделения бластомеров, что приводит к развитию двух организмов.
• Монохориальные близнецы — монозиготные близнецы, появившиеся вследствие разделения внутренней клеточной массы без разделения бластоцисты.
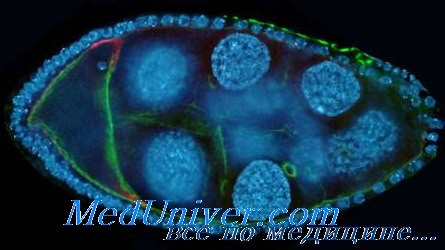
• Морула — компактный шар из примерно 16 клеток, появляющийся после первых четырех делений зиготы.
• Морфоген — вещество, производимое клетками в конкретной области эмбриона, распространяющееся из места происхождения по тканям эмбриона с образованием градиента концентрации. В зависимости от концентрации морфогена, действующего на клетку-мишень, она детерминируется к развитию по конкретной линии дифференцировки.
• Морфогенез — создание различных структур в ходе эмбриогенеза.
• Мультипотентная стволовая клетка — стволовая клетка, способная к самовоспроизводству, а также к развитию во многие типы клеток, но не в целый организм.
• Органогенез — закладка и развитие конкретных органов в ходе эмбриогенеза.
• Плод — этап развития человека с 9 нед гестации до рождения.
• Регуляционное развитие — этап развития, когда клетки еще не коммитированы, и удаление части эмбриона все равно может привести к формированию полного организма.
• Специализация — шаг на пути дифференцировки, когда клетки приобретают определенные характеристики конкретной ткани.
• Стволовая клетка — клетка, способная как к генерации другой стволовой клетки (самовоспроизводство), так и к дифференцировке в специализированные клетки в пределах ткани или целого организма.
• Тотипотентная клетка — ранняя стволовая клетка, способная к самовоспроизводству, а также к дифференцировке в любую клетку любой ткани, т.е. эмбриональные стволовые клетки тотипотентны.
• Судьба — окончательное предназначение клетки на пути развития.
• Химера — эмбрион, состоящий из двух и более клеточных мишеней, различающихся по своим генотипам. Противоположность — мозаик.
• Хорион — оболочка, развивающаяся из внешних клеток бластоцисты и формирующая впоследствии плаценту.
• Эктодерма — первичный эмбриональный листок, дающий начало нервной системе и коже.
• Эмбриогенез — развитие эмбриона.
• Эмбрион — этап развития организма человека от оплодотворения до 9 нед гестации, когда происходит разделение на плацентарные и эмбриональные ткани. В течение этого периода происходит морфогенез основных структур тела.
• Эмбриональные стволовые клетки — клетки, происходящие из внутренней клеточной массы; при подходящих условиях они могут дифференцироваться во все типы клеток и тканей эмбриона, формируя полноценный эмбрион.
• Энтодерма — первичный эмбриональный зародышевый слой, формирующий многие висцеральные органы и стенки кишечника.
• Эпибласт — наружный листок двуслойного зародыша.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Расщепление ткани в человеке что это
В норме у новорожденных при голодании количество высвобождающихся в результате протеолиза аминокислот примерно в 2-3 раза превышает подобные показатели натощак у взрослых.
Не ясны причины более высоких темпов обновления тела и разрушения мышечного белка у недоношенных новорожденных, но более высокий темп протеолиза у незрелых новорожденных может значительно влиять на нормальный рост, чтобы обеспечить достаточное количество аминокислот для ремоделирования ткани, аккреции белка и гомеостаза глюкозы.
Мало информации относительно молекулярной регуляции распада белка у новорожденных. Были проведены несколько исследований недоношенных детей с целью выяснить, какие из систем распада белка являются активными и как они функционируют. Статьи на нашем сайте дадут общие представления о катаболизме белков.
Как только белок образовался, он сразу же становится объектом распада. Некоторые белки, такие как коллаген и гемоглобин, сравнительно устойчивы к деградации, и, следовательно, их оборот происходит медленно. Другие белки распадаются легко, особенно те, которые выполняют важную регулирующую функцию либо в той или иной степени повреждены, или те, которые имеют погрешность в аминокислотной последовательности, вызванную погрешностями в транскрипции.
Детали молекулярных основ распада белка, или протеолиза, описаны не столь подробно, как система синтеза белка. Однако подобно синтезу регулирование распада белка включает компонент, направленный на специфические белки, и компонент, который регулирует общий уровень распада белка в тканях и способствует изменениям в содержании белка. Примером того, как распад отдельного белка может вызвать заболевание, является муковисцидоз (кистозный фиброз).
Удаление фенилаланина в позиции 508 из CFTR приводит к дефектам сворачивания, чувствительным к температуре и преждевременному распаду в клетке, предотвращая перемещение фенилаланина к поверхности клетки. Отсутствие CFTR в эпителиальных клетках дыхательных путей нарушает гидратацию в их просвете и увеличивает восприимчивость к инфекции. Примером согласованного увеличения скорости деградации всех белков ткани, которое позволяет всей ткани адаптироваться к изменениям окружающей среды, является усиленный протеолиз белков мышечной ткани в ответ на ряд стрессовых состояний, в том числе на голодание, ацидоз и термическую травму.
В эукариотических клетках расщепление белка осуществляется большим количеством специфических и неспецифических протеаз. Большинство этих ферментов деградации может быть связано с одной из трех основных составляющих клеточного ращепления белка: убиквитин-протеасомным путем, аутофагально-лизосомальной системой, а также кальций- или кальпаин-зависимой системой.
Убиквитин-протеасомная система в основном разрушает внутриклеточные белки, в то время как аутофагально-лизосомальная система — белки мембраны и эндоцитозные белки. Кальций-зависимые тиоловые протеазы, известные как кальпаины, широко экспрессированы и вовлечены в ряд основных клеточных процессов, хотя их физиологическая функция в развитии организма человека недостаточно понятна.
Другой класс протеолитических веществ — это семейство каспаз, или ферментов, вызывающих распад белка. Каспазы являются основными участниками апоптоза, который удаляет старые, поврежденные или потенциально опасные клетки. Исследования сигнальных путей, регулирующих распад белка, доказали, что эти процессы сложны и являются столь же тщательно контролируемыми, как и процессы синтеза белка.
Читайте также: Ткань с орнаментом полученным переплетением нитей 7 букв
Жировая ткань: роль жира в организме человека
» data-image-caption=»» data-medium-file=»https://unclinic.ru/wp-content/uploads/2020/07/zhirovaja-tkan-rol-zhira-v-organizme-cheloveka-900×600.jpg» data-large-file=»https://unclinic.ru/wp-content/uploads/2020/07/zhirovaja-tkan-rol-zhira-v-organizme-cheloveka.jpg» title=»Жировая ткань: роль жира в организме человека»>
Алена Герасимова (Dalles) Разработчик сайта, редактор
Мы много лет боремся с эпидемией ожирения, поэтому ученые уделяют все больше внимания как избыточному весу, так и ожирению, а также составу и функциям самой жировой ткани. Повышение интереса к этой теме связано с открытием в 1994 году лептина – гормона, секретируемого жировой тканью, который играет важную роль в регуляции аппетита. После этого открытия жировая ткань больше не рассматривается как энергетический резервуар, по мнению ученых – это эндокринный орган.
Что такое жировая ткань
Жировая ткань – это самый большой запас энергии в организме человека, который активируется при увеличении потребности организма в энергии. Правильное количество жира составляет 12-20% для мужчин и 20-30% для женщин.
Содержание жира у женщин физиологически выше – это совершенно нормально. С другой стороны, у тех, кто занимается спортом, могут быть более низкие значения, и это состояние не считается патологией. Следует подчеркнуть, что жировая ткань характеризуется очень высокой пластичностью и может менять свое количество в зависимости от факторов окружающей среды, рациона питания и физической активности.
Процентное содержание жира в организме
Увеличение количества жира является следствием двух процессов:
- гипертрофии – то есть увеличения размера отдельных адипоцитов;
- гиперплазии – то есть увеличения их количества.
Распределение жировой ткани непосредственно под кожей и между органами означает, что она выполняет изолирующую и амортизирующую функцию, защищая внутренние органы от механических повреждений.
Жировая структура довольно сложна. Эта гетерогенная ткань состоит из клеток, называемых адипоцитами, а также преадипоцитов, лейкоцитов, моноцитов, фибробластов, макрофагов, эндотелиальных клеток и стволовых клеток типа SVF – стромальная сосудистая фракция.
Как уже упоминалось, жировая ткань является активным эндокринным органом, который выделяет много разных веществ. Эти вещества, называемые адипокинами, выполняют разные биологические функции в организме человека:
- регулирование чувства голода и сытости;
- поддержание надлежащего энергетического баланса;
- регуляция метаболизма углеводов и жиров;
- обеспечение функционирования эндотелия сосудов;
- влияние на воспалительные и иммунные процессы.
У пациентов с лишним весом профиль секреции этих веществ изменяется, что, в свою очередь, инициирует патологические процессы, включая ожирение, резистентность к инсулину, развитие диабета и сердечно-сосудистые заболевания.
Жировая ткань в организме
Типы жировой ткани – белая и коричневая жировая ткань
Исследования показывают, что в организме человека есть 3 типа жира:
- белая жировая ткань (WAT);
- коричневая жировая (BAT);
- розовая жировая ткань – образуется из подкожного жира у беременных и при кормлении грудью.
Кроме того, выделяются бежевые адипоциты. Такие клетки считаются переходной формой между белыми и коричневыми клетками жировой ткани.
Структура отдельных типов жировой ткани различна, и это, в свою очередь, определяет, какую роль они играют в организме человека.
Название «коричневая» жировая ткань указывает на характерную окраску многочисленных митохондрий, содержащихся в ней. Эта ткань есть только у млекопитающих, и ее основная задача – поддержание постоянной температуры тела и использование избыточной энергии. Она отличается от белой жировой ткани наличием разобщенных белковых клеток белка-1 (UCP-1), так называемого термогенина, который позволяет расходовать энергию (рассеивание энергии в виде тепла).
В этом основное отличие, поскольку белая жировая ткань накапливает энергию в форме триглицеридов, которые затем можно использовать в качестве источника энергии при повышенных расходах энергии. Белые адипоциты могут эффективно выполнять эту функцию благодаря большой способности увеличивать размер. Оказывается, клетки белой жировой ткани могут увеличивать свой диаметр до 20 раз.
Увеличение количества коричневого жира наблюдается в условиях пониженной температуры окружающей среды. Долгое время считалось, что этот тип тканей встречается только у новорожденных. Тем не менее в последних исследованиях этот вид жира также был обнаружен у взрослого человека. Он расположен в основном вокруг шеи и между лопатками.
Похожее
Вы должны авторизоваться чтобы опубликовать комментарий.
Научная электронная библиотека

Колосов А. Е., Жданова О. Б., Мартусевич А. К., Ашихмин С. П.,
1.3. Краткая характеристика азотистого обмена в организме человека и млекопитающих
Азотистый обмен (АО) – совокупность химических превращений, реакций синтеза и распада азотистых соединений в организме; составная часть обмена веществ и энергии. Понятие «азотистый обмен» включает в себя белковый обмен (совокупность химических превращений в организме белков и продуктов их метаболизма), а также обмен пептидов, аминокислот, нуклеиновых кислот, нуклеотидов, азотистых оснований, азотсодержащих липидов, витаминов, гормонов и других соединений, содержащих азот.
Организм животных и человека усвояемый азот получает с пищей, в которой основным источником азотистых соединений являются белки животного и растительного происхождения. Главным фактором поддержания азотистого равновесия – состояния АО, при котором количество вводимого и выводимого азота одинаково, – служит адекватное поступление белка с пищей. Еще в СССР суточная норма белка в питании взрослого человека принята равной 100 г (или 16 г азота белка) при расходе энергии 2500 ккал.
Азотистый баланс (разность между количеством азота, который попадает в организм с пищей, и количеством азота, выводимого из организма с мочой, калом, потом) является показателем интенсивности АО в организме. Голодание или недостаточное по азоту питание приводят к отрицательному азотистому балансу, или азотистому дефициту, при котором количество азота, выводимого из организма, превышает количество азота, поступающего в организм с пищей. Положительный азотистый баланс, при котором вводимое с пищей количество азота превышает количество азота, выводимое из организма, наблюдается в период роста организма, при процессах регенерации тканей и т.д. Состояние АО в значительной степени зависит от качества пищевого белка, которое, в свою очередь, определяется его аминокислотным составом и прежде всего наличием незаменимых аминокислот.
Принято считать, что у человека и позвоночных животных АО начинается с переваривания азотистых соединений пищи в желудочно-кишечном тракте. В желудке происходит расщепление белков при участии пищеварительных протеолитических ферментов Трипсина и гастриксина с образованием полипептидов, олигопептидов и отдельных аминокислот. Из желудка пищевая масса поступает в двенадцатиперстную кишку и нижележащие отделы тонкой кишки, где пептиды подвергаются дальнейшему расщеплению, катализируемому ферментами сока поджелудочной железы трипсином, химотрипсином и карбоксипептидазой и ферментами кишечного сока аминопептидазами и дипептидазами. Наряду с пептидами, в тонкой кишке расщепляются сложные белки (например, нуклеопротеины) и нуклеиновые кислоты. Существенный вклад в расщепление азотсодержащих биополимеров вносит и микрофлора кишечника. Олигопептиды, аминокислоты, нуклеотиды, нуклеозиды и др. всасываются в тонкой кишке, поступают в кровь и с ней разносятся по всему организму. Белки тканей организма в процессе постоянного обновления также подвергаются протеолизу под действием тканевых протеаз (пептидаз и катепсинов), а продукты распада тканевых белков попадают в кровь. Аминокислоты могут быть использованы для нового синтеза белков и других соединений (пуриновых и пиримидиновых оснований, нуклеотидов, порфиринов и т.д.), для получения энергии (например, посредством включения в цикл трикарбоновых кислот) или могут быть подвергнуты дальнейшей деградации с образованием конечных продуктов АО, подлежащих выведению из организма.
Аминокислоты, поступающие в составе белков пищи, используются для синтеза белков органов и тканей организма. Они участвуют также в образовании многих других важных биологических соединений: пуриновых нуклеотидов (глутамин, глицин, аспарагиновая кислота) и пиримидиновых нуклеотидов (глутамин, аспарагиновая кислота), серотонина (триптофан), меланина (фенилалпнин, тирозин), гистамина (гистидин), адреналина, норадреналина, тирамина (тирозин), полиаминов (аргинин, метионин), холина (метионин), порфиринов (глицин), креатина (глицин, аргинин, метионин), коферментов, сахаров и полисахаридов, липидов и т.д. Важнейшей для организма химической реакцией, в которой участвуют практически все аминокислоты, является трансаминирование, заключающееся в обратимом ферментативном переносе ?-аминогруппы аминокислот на ?-углеродный атом кетокислот или альдегидов. Трансаминирование является принципиальной реакцией биосинтеза заменимых аминокислот в организме. Активность ферментов, катализирующих реакции трансаминирования, – аминотрансфераз – имеет большое клинико-диагностическое значение.
Читайте также: Чайка 142м не шьет тонкую ткань
Деградация аминокислот может протекать по нескольким различным путям. Большинство аминокислот способно подвергаться декарбоксилированию при участии ферментов декарбоксилаз с образованием первичных аминов, которые затем могут окисляться в реакциях, катализируемых моноаминоксидазой или диаминоксидазой. При окислении биогенных аминов (гистамина, серотонина, тирамина, ?-аминомасляной кислоты) оксидазами образуются альдегиды, подвергающиеся дальнейшим превращениям, и аммиак, основным путем дальнейшего метаболизма которого является образование мочевины.
Другим принципиальным путем деградации аминокислот является окислительное дезаминирование с образованием аммиака и кетокислот. Прямое дезаминирование L-аминокислот в организме животных и человека протекает крайне медленно, за исключением глутаминовой кислоты, которая интенсивно дезаминируется при участии специфического фермента глутаматдегидрогеназы. Предварительное трансаминирование почти всех ?-аминокислот и дальнейшее дезаминирование образовавшейся глутаминовой кислоты на ?-кетоглутаровую кислоту и аммиак является основным механизмом дезаминирования природных аминокислот.
Продуктом разных путей деградации аминокислот является аммиак, который может образовываться и в результате метаболизма других азотсодержащих соединений (например, при дезаминировании аденина, входящего в состав никотинамидадениндинуклеотида – НАД). Основным путем связывания и нейтрализации токсичного аммиака у уреотелических животных (животные, у которых конечным продуктом АО, является мочевина) служит так называемый цикл мочевины (синоним: орнитиновый цикл, цикл Кребса-Гензелейта), протекающий в печени. Он представляет собой циклическую последовательность ферментативных реакций, в результате которой из молекулы аммиака или амидного азота глутамина, аминогруппы аспарагановой кислоты и диоксида углерода осуществляется синтез мочевины. При ежедневном потреблении 100 г белка суточное выведение мочевины из организма составляет около 30 г. У человека и высших животных существует еще один путь нейтрализации аммиака – синтез амидов дикарбоновых кислот аспарагана и глутамина из соответствующих аминокислот. У урикотелических животных (рептилии, птицы) конечным продуктом АО является мочевая кислота.
В результате расщепления нуклеиновых кислот и нуклеопротеинов в желудочно-кишечном тракте образуются нуклеотиды и нуклеозиды. Олиго- и моно-нуклеотиды при участии различных ферментов (эстераз, нуклеотидаз, нуклеозидаз, фосфорилаз) превращаются затем в свободные пуриновые и пиримидиновые основания.
Дальнейший путь деградации пуриновых оснований аденина и гуанина состоит в их гидролитическом дезаминировании под влиянием ферментов аденазы и гуаназы с образованием соответственно гипоксантина (6-оксипурина) и ксантина (2,6-диоксипурина), которые затем превращаются в мочевую кислоту в реакциях, катализируемых ксантиноксидазой. Мочевая кислота – один из конечных продуктов АО и конечный продукт обмена пуринов у человека – выводится из организма с мочой. У большинства млекопитающих имеется фермент уриказа, который катализирует превращение мочевой кислоты в экскретируемый аллантоин.
Деградация пиримидиновых оснований (урацила, тимина) состоит в их восстановлении с образованием дигидропроизводных и последующем гидролизе, в результате которого из урацила образуется ?-уреидопропионовая кислота, а из нее – аммиак, диоксид углерода и ?-аланин, а из тимина – ?-аминоизомасляная кислота, диоксид углерода и аммиак. Диоксид углерода и аммиак могут далее включаться в мочевину через цикл мочевины, а ?-аланин участвует в синтезе важнейших биологически активных соединений – гистидинсодержащих дипептидов карнозина (?-аланил-L-гистидина) и анзерина (?-аланил-N-метил-L-гистидина), обнаруживаемых в составе экстрактивных веществ скелетных мышц, а также в синтезе пантотеновой кислоты и кофермента А.
Таким образом, разнообразные превращения важнейших азотистых соединений организма связаны между собой в единый обмен. Сложный процесс АО регулируется на молекулярном, клеточном и тканевом уровнях. Регуляция АО в целом организме направлена на приспособление интенсивности АО к изменяющимся условиям окружающей и внутренней среды и осуществляется нервной системой как непосредственно, так и путем воздействия на железы внутренней секреции.
У здоровых взрослых людей содержание азотистых соединений в органах, тканях, биологических жидкостях находится на относительно постоянном уровне. Избыток азота, поступившего с пищей, выводится с мочой и калом, а при недостатке азота в пище нужды организма в нем могут покрываться за счет использования азотистых соединений тканей тела. При этом состав мочи изменяется в зависимости от особенностей АО и состояния азотистого баланса. В норме при неизменном режиме питания и относительно стабильных условиях окружающей среды из организма выделяется постоянное количество конечных продуктов АО, а развитие патологических состояний приводит к его резкому изменению. Значительные изменения экскреции азотистых соединений с мочой, в первую очередь экскреции мочевины, могут наблюдаться и при отсутствии патологии в случае существенного изменения режима питания (например, при изменении количества потребляемого белка), причем концентрация остаточного азота в крови меняется незначительно.
При исследовании АО необходимо учитывать количественный и качественный состав принимаемой пищи, количественный и качественный состав азотистых соединений, выделяемых с мочой и калом и содержащихся в крови. Для исследования АО применяют азотистые вещества, меченные радионуклидами азота, фосфора, углерода, серы, водорода, кислорода, и наблюдают за миграцией метки и включением ее в состав конечных продуктов АО Широко используют меченые аминокислоты, например 15N-глицин, которые вводят в организм с пищей или непосредственно в кровь. Значительная часть меченого азота глицина пищи выводится в составе мочевины с мочой, а другая часть метки попадает в тканевые белки и выводится из организма крайне медленно. Проведение исследования АО необходимо для диагностики многих патологических состояний и контроля эффективности лечения, а также при разработке рациональных схем питания, в т.ч. лечебного.
Патологию АО (вплоть до клинически значимой) вызывает белковая недостаточность. Ее причиной может стать общее недоедание, продолжительный дефицит белка или незаменимых аминокислот в рационе, недостаток углеводов и жиров, обеспечивающих энергией процессы биосинтеза белка в организме. Белковая недостаточность может быть обусловлена преобладанием процессов распада белков над их синтезом не только в результате алиментарного дефицита белка и других важнейших пищевых веществ, но и при тяжелой мышечной работе, травмах, воспалительных и дистрофических процессах, ишемии, инфекции, обширных ожогах, дефекте трофической функции нервной системы, недостаточности гормонов анаболического действия (гормона роста, половых гормонов, инсулина), избыточном синтезе или избыточном поступлении извне стероидных гормонов и т.п. Нарушение усвоения белка при патологии желудочно-кишечного тракта (ускоренная эвакуация пищи из желудка, гипо- и анацидные состояния, закупорка выводного протока поджелудочной железы, ослабление секреторной функции и усиление моторики тонкой кишки при энтеритах и энтероколитах, нарушение процесса всасывания в тонкой кишке и др.) также может приводить к белковой недостаточности. Белковая недостаточность ведет к дискоординации АО и характеризуется резко выраженным отрицательным азотистым балансом.
Известны случаи нарушения синтеза определенных белков, а также генетически обусловленного синтеза аномальных белков, например при гемоглобинопатиях, миеломной болезни и др.
Патология АО, заключающаяся в нарушении обмена аминокислот, часто связана с аномалиями процесса трансаминирования: уменьшением активности аминотрансфераз при гипо- или авитаминозах В6, нарушением синтеза этих ферментов, недостатком кетокислот для трансаминирования в связи с угнетением цикла трикарбоновых кислот при гипоксии и сахарном диабете и т.д. Снижение интенсивности трансаминирования приводит к угнетению дезаминирования глутаминовой кислоты, а оно, в свою очередь, – к повышению доли азота аминокислот в составе остаточного азота крови (гипераминоацидемии), общей гиперазотемии и аминоацидурии. Гипераминоацидемия, аминоацидурия и общая азотемия характерны для многих видов патологии АО. При обширных поражениях печени и других состояниях, связанных с массивным распадом белка в организме, нарушаются процессы дезаминирования аминокислот и образования мочевины таким образом, что возрастают концентрация остаточного азота и содержание в нем азота аминокислот на фоне снижения относительного содержания в остаточном азоте азота мочевины (так называемая продукционная азотемия). Продукционная азотемия, как правило, сопровождается выведением избытка аминокислот с мочой, поскольку даже в случае нормального функционирования почек фильтрация аминокислот в почечных клубочках происходит интенсивнее, чем их реабсорбция в канальцах. Заболевания почек, обтурация мочевых путей, нарушение почечного кровообращения приводят к развитию ретенционной азотемии, сопровождающейся нарастанием концентрации остаточного азота в крови за счет повышения содержания в крови мочевины. Обширные раны, тяжелые ожоги, инфекции, повреждения трубчатых костей, спинного и головного мозга, гипотиреоз, болезнь Иценко–Кушинга и многие другие тяжелые заболевания сопровождаются аминоацидурией. Она характерна и для патологических состояний, протекающих с нарушением процессов реабсорбции в почечных канальцах: болезни Вильсона–Коновалова (гепатоцеребральная дистрофия), нефронофтизе Фанкони и др. Эти болезни относятся к многочисленным генетически обусловленным нарушениям АО. Избирательное нарушение реабсорбции цистина и цистинурия с генерализованным нарушением обмена цистина на фоне общей аминоацидурии сопровождает так называемый цистиноз. При этом заболевании кристаллы цистина откладываются в клетках ретикулоэндотелиальной системы. Наследственное заболевание фенилкетонурия характеризуется нарушением превращения фенилаланина в тирозин в результате генетически обусловленной недостаточности фермента фенилала-
нин – 4-гидроксилазы, что вызывает накопление в крови и моче непревращенного фенилаланина и продуктов его обмена – фенилпировиноградной и фенилуксусной кислот. Нарушение превращений этих соединений характерно и для вирусного гепатита.
Читайте также: Seda ткань что это такое
Тирозинемию, тирозинурию и тирозиноз отмечают при лейкозах, диффузных заболеваниях соединительной ткани (коллагенозах) и других патологических состояниях. Они развиваются вследствие нарушения трансаминирования тирозина. Врожденная аномалия окислительных превращений тирозина лежит в основе алкаптонурии, при которой в моче накапливается непревращенный метаболит этой аминокислоты – гомогентизиновая кислота. Нарушения пигментного обмена при гипокортицизме связаны с угнетением превращения тирозина в меланин вследствие ингибирования фермента тирозиназы (полное выпадение синтеза этого пигмента характерно для врожденной аномалии пигментации – альбинизма).
При хроническом гепатите, сахарном диабете, остром лейкозе, хроническом миело- и лимфолейкозе, лимфогранулематозе, ревматизме и склеродермии нарушается обмен триптофана и его метаболиты 3-оксикинуренин, ксантуреновая и 3-оксиантраниловая кислоты, обладающие токсическими свойствами, накапливаются в крови. К патологии АО относятся и состояния, связанные с нарушением выделения почками креатинина и накоплением его в крови. Усиление экскреции креатинина сопровождает гиперфункцию щитовидной железы, а снижение экскреции креатинина при повышенном выведении креатина – гипотиреоз.
При массивном распаде клеточных структур (голодание, тяжелая мышечная работа, инфекции и др.) отмечают патологическое нарастание концентрации остаточного азота за счет увеличения относительного содержания в ней азота мочевой кислоты (в норме концентрация мочевой кислоты в крови не превышает 0,4 ммоль/л).
В пожилом возрасте снижаются интенсивность и объем синтеза белка за счет непосредственного угнетения биосинтетической функции организма и ослабления его способности усваивать аминокислоты пищи; развивается отрицательный азотистый баланс. Нарушения обмена пуринов у людей пожилого возраста приводят к накоплению и отложению в мышцах, суставах и хрящах солей мочевой кислоты – уратов. Коррекция нарушений АО в пожилом возрасте может быть осуществлена за счет специальных диет, содержащих полноценные животные белки, витамины и микроэлементы, с ограниченным содержанием пуринов.
Азотистый обмен у детей отличается рядом особенностей, в частности положительным азотистым балансом как необходимым условием роста. Интенсивность процессов АО на протяжении роста ребенка подвергается изменениям, особенно ярко выраженным у новорожденных и детей раннего возраста. В течение первых 3-х дней жизни азотистый баланс отрицателен, что объясняется недостаточным поступлением белка с пищей. В этот период обнаруживается транзиторное повышение концентрации остаточного азота в крови (так называемая физиологическая азотемия), иногда достигающее 70 ммоль/л; к концу 2-й нед. жизни концентрация остаточного азота снижается до уровня, отмечаемого у взрослых. Количество выделяемого почками азота нарастает в течение первых 3-х дней жизни, после чего снижается и вновь начинает увеличиваться со 2-й недели жизни параллельно возрастающему количеству пищи.
Наиболее высокая усвояемость азота в организме ребенка наблюдается у детей первых месяцев жизни. Азотистый баланс заметно приближается к равновесию в первые 3-6 мес. жизни, хотя и остается положительным. Интенсивность белкового обмена у детей достаточно высока у детей 1-го года жизни обновляется около 0,9 г белка на 1 кг массы тела в сутки, в 1-3 года –
0,8 г/кг/сут, у детей дошкольного и школьного возраста – 0,7 г/кг/сут.
Средние величины потребности в незаменимых аминокислотах, по данным ФАО ВОЗ (1985), у детей в 6 раз больше, чем у взрослых (незаменимой аминокислотой для детей в возрасте до 3 месяцев является цистин, а до 5 лет – и гистидин). Более активно, чем у взрослых, у детей протекают процессы трансаминирования аминокислот. Однако в первые дни жизни у новорожденных из-за относительно низкой активности некоторых ферментов отмечаются гипераминоацидемия и физиологическая аминоацидурия в результате функциональной незрелости почек. У недоношенных, кроме того, имеет место аминоацидурия перегрузочного типа, т.к. содержание свободных аминокислот в плазме их крови выше, чем у доношенных детей. На первой неделе жизни азот аминокислот составляет 3-4 % общего азота мочи (по некоторым данным – до 10 %), и лишь к концу 1-го года жизни его относительное содержание снижается до 1 %. У детей 1-го года жизни выведение аминокислот в расчете на 1 кг массы тела достигает величин выведения их у взрослого человека, экскреция азота аминокислот, достигающая у новорожденных 10 мг/кг массы тела, на 2-м году жизни редко превышает 2 мг/кг массы тела. В моче новорожденных повышено (по сравнению с мочой взрослого человека) содержание таурина, треонина, серина, глицина, аланина, цистина, лейцина, тирозина, фенилаланина и лизина. В первые месяцы жизни в моче ребенка обнаруживаются также этаноламин и гомоцитруллин. В моче детей 1-го года жизни преобладают аминокислоты пролин и [гидр]оксипролин.
Исследования важнейших азотистых компонентов мочи у детей показали, что соотношение мочевой кислоты, мочевины и аммиака в процессе роста существенно изменяется. Так, первые 3 мес. жизни характеризуются наименьшим содержанием в моче мочевины (в 2–3 раза меньше, чем у взрослых) и наибольшей экскрецией мочевой кислоты. Дети в первые три месяца жизни выделяют 28,3 мг/кг массы тела мочевой кислоты, а взрослые – 8,7 мг/кг. Относительно высокая экскреция у детей первых месяцев жизни мочевой кислоты способствует иногда развитию мочекислого инфаркта почек. Количество мочевины в моче нарастает у детей в возрасте от 3 до 6 месяцев, а содержание мочевой кислоты в это время снижается. Содержание аммиака в моче детей в первые дни жизни невелико, но затем резко возрастает и держится на высоком уровне на протяжении всего 1-го года жизни.
Характерной особенностью АО у детей является физиологическая креатинурия. Креатин обнаруживается еще в амниотической жидкости; в моче он определяется в количествах, превышающих содержание креатина в моче взрослых, начиная с периода новорожденности и до периода полового созревания. Суточная экскреция креатинина (дегидроксилированного креатина) с возрастом увеличивается, в то же время по мере нарастания массы тела ребенка относительное содержание азота креатинина мочи снижается. Количество креатинина, выводимого с мочой за сутки, у доношенных новорожденных составляет 10-13 мг/кг, у недоношенных – 3 мг/кг, у взрослых не превышает 30 мг/кг.
При выявлении в семье врожденного нарушения АО необходимо проведение медико-генетического анализа.
1. Березов Т.Т., Коровкин Б.Ф. Биологическая химия. М., 1982. – С. 431.
2. Вельтищев Ю.Е. с соавт. Обмен веществ у детей. М., 1983. – С. 53,
3. Дудел Дж. с соавт. Физиология человека / пер. с англ. – М., 1985. – т. 1-4.
4. Зилва Дж.Ф., Пэннелл П.Р. Клиническая химия в диагностике и лечении / пер. с англ. – М., 1988. – С. 298-398.
5. Кон Р.М., Рой К.С. Ранняя диагностика болезней обмена веществ / пер. с англ. – М., 1986. – С. 211.
6. Лабораторные методы исследования в клинике / под ред. В.В. Меньшикова. – М., 1987. – С. 222.
7. Ленинджер А. Основы биохимии / пер. с англ. М., 1985. – Т. 2.
8. Мазурин А.В., Воронцов И.М. Пропедевтика детских болезней. М., 1985. – С. 322.
9. Руководство по педиатрии / под. ред. У.Е. Бермана и В.К. Вогана: пер. с англ. – М., 1987. – кн. 2. – С. 337
10. Страйер Л. Биохимия / пер. с англ. – М., 1985. – т. 2. – С. 233.