
Известно, что фосфоролитический распад играет ключевую роль в мобилизации полисахаридов.
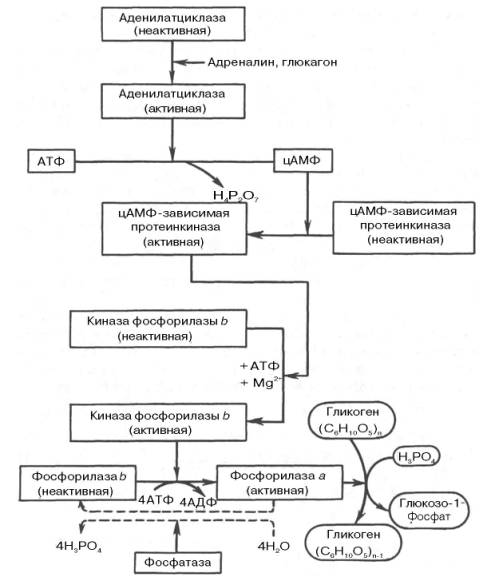
Рис. 10.1. Гормональная регуляция фосфоролитического отщепления остатка глюкозы от гликогена.
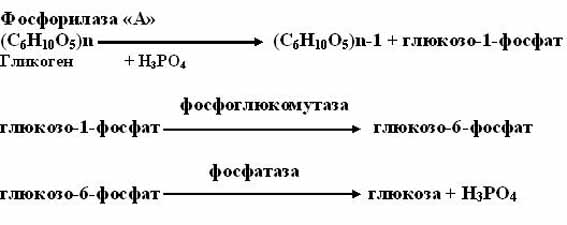
Фосфорилазы переводят полисахариды (в частности, гликоген) из запасной формы в метаболически активную форму; в присутствии фосфо-рилазы гликоген распадается с образованием фосфорного эфира глюкозы (глюкозо-1-фосфата) без предварительного расщепления на более крупные обломки молекулы полисахарида. В общей форме эту реакцию можно представить в следующем виде:
где (С6Н10О5)n означает полисахаридную цепь гликогена, а (С6Н10О5)n-1,– ту же цепь, но укороченную на один глюкозный остаток.
На рис. 10.1 изображены процесс распада гликогена до глюкозо-1-фосфата и участие в этом процессе цАМФ. Фермент фосфорилаза существует в двух формах, одна из которых (фосфорилаза а) активна, в то время как другая (фосфорилаза b) обычно неактивна. Обе формы могут диссоциировать на субъединицы. Фосфорилаза b состоит из двух субъединиц, а фосфорилаза а – из четырех. Превращение фосфо-рилазы b в фосфорилазу а осуществляется фосфорилированием белка:
2 Фосфорилаза b + 4 АТФ –> Фосфорилаза а + 4 АДФ.
Катализируется эта реакция ферментом, который называется киназой фосфорилазы b. Установлено, что эта киназа может существовать как в активной, так и в неактивной форме. Неактивная киназа фосфорилазы превращается в активную под влиянием фермента протеинкиназы (киназа киназы фосфорилазы), и не просто протеинкиназы, а цАМФ-зависимой протеинкиназы.
Активная форма последней образуется при участии цАМФ, которая в свою очередь образуется из АТФ под действием фермента аденилатцик-лазы, стимулируемой, в частности, адреналином и глюкагоном. Увеличение содержания адреналина в крови приводит в этой сложной цепи реакций к превращению фосфорилазы b в фосфорилазу а и, следовательно, к освобождению глюкозы в виде глюкозо-1-фосфата из запасного полисахарида гликогена. Обратное превращение фосфорилазы а в фосфорилазу b катализируется ферментом фосфатазой (эта реакция практически необратима).
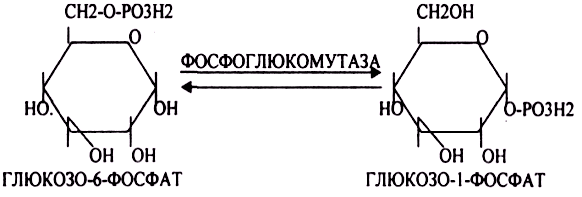
Образовавшийся в результате фосфоролитического распада гликогена глюкозо-1-фосфат превращается под действием фосфоглюкомутазы в глюкозо-6-фосфат. Для осуществления данной реакции необходима фосфо-рилированная форма фосфоглюкомутазы, т.е. ее активная форма, которая образуется, как отмечалось, в присутствии глюкозо-1,6-бисфосфата .

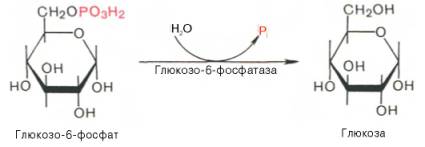
Образование свободной глюкозы из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы. Данный фермент катализирует гидролитическое отщепление фосфата:
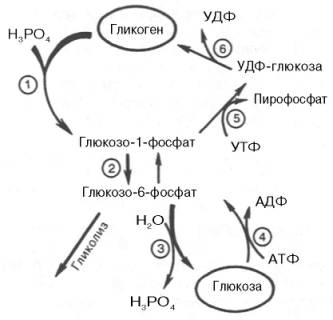
Жирными стрелками указан путь распада, тонкими — путь синтеза. Цифрами обозначены ферменты: 1 — фосфорилаза; 2 — фос-фоглюкомутаза; 3 — глюкозо-6-фосфатаза; 4 — гексокиназа (глюкокиназа); 5 — глюко-зо-1-фосфат-уридилтрансфераза; 6 — глико-генсинтаза.
Заметим, что фосфорилированная глюкоза в противоположность неэте-рифицированной глюкозе не может легко диффундировать из клеток. Печень содержит гидролитический фермент глюкозо-6-фосфатазу, который и обеспечивает возможность быстрого выхода глюкозы из этого органа. В мышечной ткани глюкозо-6-фосфатаза практически отсутствует.
На рис. 10.2 отражены представления о путях распада и синтеза гликогена в печени.
Можно считать, что сохранение постоянства концентрации глюкозы в крови является результатом одновременного протекания двух процессов: поступления глюкозы в кровь из печени и потребления ее из крови тканями, где она используется в первую очередь как энергетический материал.
В тканях (в том числе в печени) распад глюкозы происходит двумя основными путями: анаэробным (при отсутствии кислорода) и аэробным, для осуществления которого необходим кислород.
Биосинтез и распад гликогена
БИОСИНТЕЗ И РАСПАД ГЛИКОГЕНА В ТКАНЯХ.
Было установлено, что гликоген может синтезироваться практически во всех органах и тканях. Однако наибольшая его концентрация обнаружена в печени (2-6%) и мышцах (0,5-2%). Поскольку мышечная масса организма человека велика, то большая часть гликогена организма содержится в мышцах. Глюкоза из крови легко проникает в клетки органов и тканей, проходя через биологические мембраны клеток. Как только глюкоза поступает в клетку, она метаболизируется в ней в результате первой химической реакции. фосфорилирование глюкозы происходит в присутствии АТФ и фермента — гексокиназы. Глюкоза превращается в глюкозо-6-фосфат . Этот эфир глюкозы теперь будет использоваться в анаболических и катаболических реакциях. Глюкоза из клетки может выйти только после реакции гидролиза при участии глюкозо-6-фосфатазы. Этот фермент есть в печени, почках, эпителии кишечника. В других органах, тканях его нет. Процесс биосинтеза гликогена протекает в 4 стадии:
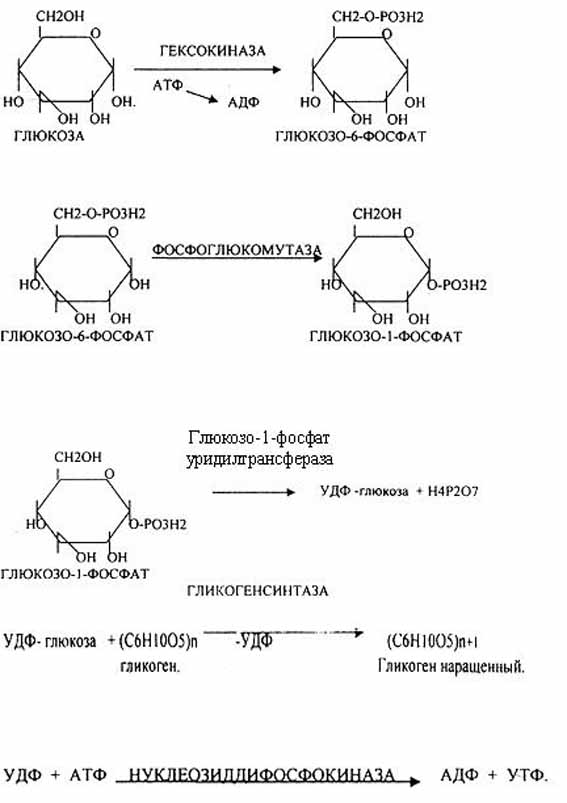
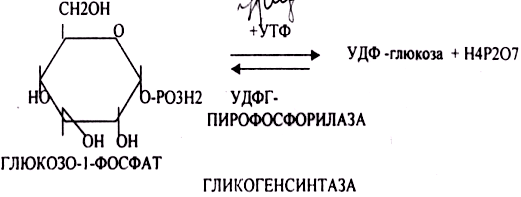
Гликогенсинтаза – является трансферазой, которая переносит остатки глюкозы, входящие в УДФ- глюкозу, на гликозидную связь остаточного в клетке гликогена, при этом образуется альфа(1,4)-гликозидные связи.
Образование альфа(1,6)-гликозидных связей в точках ветвления гликогена катализирует специальный гликоген-ветвящий фермент.
Образовавшийся в последней реакции УДФ, превращается в УТФ, при этом при биосинтезе гликогена поглощается еще 1 молекула АТФ.
Таким образом, на каждую молекулу глюкозы, включающуюся в структуру гликогена, расходуется 2 молекулы АТФ.
Гликоген в клетках накапливается во время пищеварения и рассматривается как резервная форма глюкозы, которая используется клетками в промежутках между приёмами пищи.
Читайте также: Ткань полиэстер 100 нити или волокна
Существуют 2 пути распада гликогена в тканях:
1. фосфоролитический путь (основной путь)
Протекает в печени, почках, эпителии кишечника. Схематически его можно записать в виде 3 реакций:
2. амилолитический путь (неосновной).
Протекает в печени при участии 3 ферментов: альфа -амилазы, амило-1,6-гликозидазы, гамма — амилазы.
Альфа – амилаза расщепляет в структуре крахмала альфа-1,4-гликозидные связи, амило-1,6-гликозидаза-гликозидные связи в точках ветвления, гамма-амилаза гидролизует концевые гликозидные связи в боковых ветвях гликогена.
ГЛИКОГЕНОЗЫ — болезни, связанные с нарушением процессов распада гликогена, при этом в клетках печени, почек, мышц гликоген накапливается в большом количестве. Клинически эти заболевания проявляются увеличением печени, мышечной слабостью, гипоглюкоземией натощак. Больные умирают в раннем детском возрасте. Наиболее часто встречаются следующие заболевания:
1.Болезнь Герса (генетический дефект фермента — фосфорилазы печени ).
2. Болезнь Мак-Ардля (генетический дефект фермента — фосфорилазы мышц ).
3. Болезнь Помпе (генетический дефект фермента — амило- 1,4-гликозидазы ).
4. Болезнь Кори (генетический дефект фермента — амило-1.6-гликозидазы ).
5. Болезнь Гирке (генетический дефект фермента — глюкозо-
АГЛИКОГЕНОЗЫ- болезни связанные с нарушением процессов синтеза гликогена в тканях. Характерными проявлениями нарушения синтеза гликогена являются: резкая гипогликемия натощак, рвоты, судороги, потеря сознания. Углеводное голодание клеток мозга приводит к нарушению психо-физического развития у детей. Смерть наступает в раннем детском возрасте. Наиболее часто встречаются:
1. Болезнь Льюиса (генетический дефект фермента – гликогенсинтазы ).
2. Болезнь Андерсена ( генетический дефект фермента — гликоген-ветвящего ).
37. Биосинтез и распад гликогена в тканях. Биологическая роль этих процессов. Гликогеновые болезни.
Гликоген может синтезироваться практически во всех органах и тканях. Большая часть гликогена содержится в мышцах.
Глюкоза из крови легко проникает в клетки органов и тканей, проходя через биологические мембраны клеток. Как только глюкоза заходит в клетку, она превращается в глюкозо-6-фосфат. Процесс биосинтеза протекает в 4 стадии:


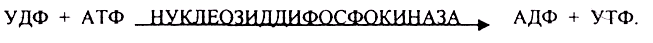
ГЛИКОГЕНСИНТАЗА — ТРАНСФЕРАЗА, которая переносит остатки глюкозы, входящие в УДФ- глюкозу, на ГЛИКОЗИДНУЮ связь остаточного в клетке гликогена. Образование альфа(1,6)-ГЛИКОЗИДНЫХ связей катализирует специальный ГЛИКОГЕН-ВЕТВЯЩИЙ фермент. Образовавшийся в последней реакции УДФ, превращается в УТФ, при этом поглощается I молекула АТФ. Гликоген в клетках накапливается во время пищеварения и рассматривается как резервная форма глюкозы, которая используется клетками в промежутках между приёмами пищи.
РАСПАД ГЛИКОГЕНА. Может идти 2 путями:
1. ОСНОВНОЙ ФОСФОРОЛИТИЧЕСКИЙ ПУТЬ
Он протекает в печени, почках, эпителии кишечника. Схематично можно записать в виде 3 реакций.

2.НЕОСНОВНОЙ АМИЛОЛИТИЧЕСКИЙ. Его доля незначительна. Протекает в печени при участии 3 ферментов: альфа — АМИЛАЗА, АМИЛО-1,6-ГЛИКОЗИДАЗА, гамма -АМИЛАЗА.
Это наследственное нарушение обмена гликогена, которое связано с недостатком какого-либо из ферментов, участвующих в синтезе или распаде гликогена. ГЛИКОГЕНОЗЫ — болезни, связанные с нарушением процессов распада гликогена. Клинически проявляются увеличением печени, мышечной слабостью, ГИПОГЛЮКОЗЭМИЕЙ натощак. Наиболее часто встречаются:
1.болезнь ФЕРСА — ФОСФОРИЛАЗА ПЕЧЕНИ.
2. Болезнь МАК-АРДЛЯ — ФОСФОРИЛАЗА МЫШЦ.
3. Болезнь ПОМПЕ- альфа- 1,4-ГЛИКОЗИДАЗА.
4. Болезнь КОРИ — АМИЛО-1.6-ГЛИКОЗИДАЗА.
5. Болезнь ГИРКЕ — ГЛЮКОЗО-6-ФОСФОТАЗА.
АГЛИКОГЕНОЗЫ характеризуются признаками нарушения синтеза. Клинически проявляются резкой ГИПОГЛЮКОЗЭМИЕЙ натощак, рвотой, судорогами, потерей сознания. Наиболее часто встречаются:
1. болезнь ЛЬЮИСА — ГЛИКОГЕНСИНТЕТАЗА.
2. Болезнь АНДЕРСЕНА — ГЛИКОГЕН-ВЕТВЯЩИЙ фермент.
38. Пути образования глюкозы в организме. Глюконеогенез. Возможные предшественники, последовательность реакций, биологическая роль.
ГЛЮКОНЕОГЕНЕЗ — это биосинтез глюкозы из неуглеводных предшественников, главными из которых являются ПИРУВАТ, ЛАКТАТ, ГЛИЦЕРИН, ряд АК, тем или иным путем превращающиеся в ФУМАРАТ, который в дальнейшем превращается в ЩУК. Другие АК превращаются в ПИРУВАТ. Главным местом синтеза глюкозы является печень. ГЛЮКОНЕОГЕНЕЗ обеспечивает синтез глюкозы, а также возврат ЛАКТАТА, образованного в реакциях анаэробного ГЛИКОЛИЗА, в клеточный фонд углеводов. За счет этого процесса поддерживается уровень глюкозы при углеводном голодании, сахарном диабете. Большинство реакций ГЛЮКОНЕОГЕНЕЗА представляют собой обратные реакции ГЛИКОЛИЗУ, за исключением трёх (ГЕКСОКИНАЗНОЙ, ФОСФОФРУКТОКИНАЗНОЙ, ПИРУВАТКИНАЗНОЙ), которые при ГЛюКОНЕОГЕНЕЗЕ имеют обходные пути.
Первая обходная реакция ГЛЮКОНЕОГЕНЕЗА связана с образованием 2-ФОСФОЕНОЛ ПВК и протекает в две стадии:

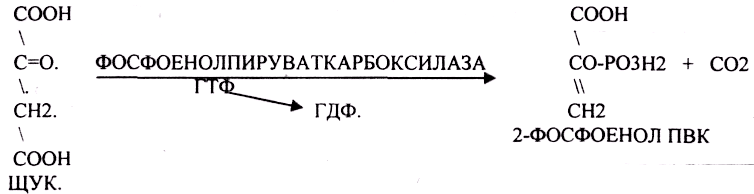
Вторая обходная реакция связана с образованием фруктозо-6-фосфата.
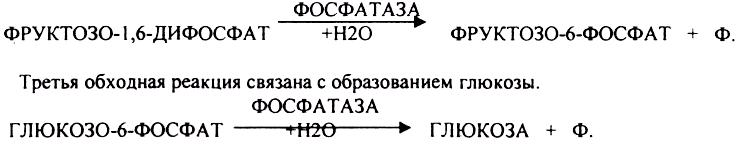
Образовавшаяся глюкоза может вновь использоваться клетками как пластический и энергетический материал, откладываться про запас в виде гликогена. В организме взрослого человека за сутки может быть образованно 80 гр. глюкозы.
Синтез и распад гликогена.
Синтез гликогена протекает не во всех тканях, а только в печени, мышцах и в лейкоцитах.
После образования глюкозо-6-фосфата (гексокиназная реакция) происходит внутримолекулярный перенос остатка фосфорной кислоты из 6-го положения в 1-е. При этом образуется глюкозо-1-фосфат:
Читайте также: Комбинирование тканей в женской одежде платье

После изомеризации глюкозо-6-фосфата в глюкозо-1-фосфат протекает дополнительная активация глюкозного фрагмента. При этом расходуется 1 молекула УТФ, что эквивалентно расходованию 1-й молекулы АТФ. В результате образуется активированная форма — УДФ-глюкоза:
Затем с УДФ глюкозный остаток переносится на молекулу гликогена. Удлинение цепи гликогена катализирует фермент гликогенсинтетаза. Таким образом, цепь гликогена становится на 1 глюкозный фрагмент длиннее. Гликоген, в отличие от растительного крахмала, более сильно разветвлен. Для формирования ответвлений существует специальный фермент, который называется «гликогенветвящий фермент» (стр.242 учебника).
Молекула гликогена синтезируется не с «нуля», а происходит постепенное удлинение уже имеющегося кусочка цепи: «затравки». И при распаде гликогена никогда не происходит полного разрушения его молекул.
Для включения одного остатка глюкозы в молекулу гликогена клетка расходует 2 молекулы АТФ. При распаде гликогена эта АТФ не регенерирует, а освобождается только Фн (неорганический фосфат).
Ключевым ферментом синтеза гликогена является гликогенсинтаза. Это «пункт вторичного контроля». Ее Vmax больше, чем у гексокиназы, но меньше, чем у любого другого фермента на пути синтеза гликогена.
Регуляция гликогенсинтазы: она активируется избытком глюкозо-6-фосфата. Поэтому если глюкозо-6-фосфат другими путями утилизируется медленно, то возрастание его концентрации приводит к увеличению скорости синтеза гликогена.
Реакция, катализируемая гликогенсинтазой, необратима.
В определенных условиях гликоген способен распадаться. Для этого существует свой обходной обратный путь. Его ключевым ферментом является гликогенфосфорилаза (фосфорилаза). Этот фермент расщепляет молекулу гликогена с участием Фн до глюкозо-1-фосфата и гликогена, укороченного на один глюкозный фрагмент: (С6Н10О5)n + H3PO4 —-> (C6H10O5)n-1 + глюкозо-1-фосфат
Фосфорилаза — ключевой (то есть лимитирующий и регуляторный) фермент распада гликогена.
Регуляция гликогенфосфорилазы: угнетается избытком АТФ, активируется избытком АДФ.
Г б ф — п у т ь. (гексозобисфосфатный путь распада углеводов)
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ГБФ-ПУТИ.
1. Это главный путь распада углеводов до конечных продуктов. Во многих клетках это — единственный путь. Так распадается 70-75% глюкозы, которая поступает в клетку.
2. Только ГБФ-путь дает клетке энергию в виде АТФ. Это основной источник получения энергии в клетке.
3. Это самый длинный путь распада углеводов.
1-й этап протекает в цитоплазме, дает 8 молекул АТФ при распаде 1 молекулы глюкозы или 9АТФ при распаде одного глюкозного фрагмента гликогена. Заканчивается образованием 2-х молекул пирувата (ПВК).
2-й и 3-й этапы — (исключительно аэробные!) в митохондриях с обязательным участием кислорода, дают 30 АТФ в расчете на одну молекулу глюкозы.
2-й этап ГБФ-пути называется «окислительное декарбоксилирование пирувата» и катализируется пируватдегидрогеназным комплексом (смотрите лекции «Биологическое окисление» — удлиненная цепь митохондриального окисления). На 2-м этапе от молекулы ПВК отнимаются два атома водорода, и пируват превращается в Ацетил-кофермент А (АцКоА), одновременно происходит отщепление СО2. Два атома водорода идут на НАД, а затем по цепи митохондриального окисления передаются на О2 с образованием Н2О и 3 молекул АТФ. Поэтому в расчете на одну молекулу исходной глюкозы 2-й этап дает 6 АТФ.
В 3-й этап вступает молекула АцетилКоА, который образуется в результате 2-го этапа. Этот 3-й этап называется циклом трикарбоновых кислот (ЦТК) (смотрите лекции “Митохондриальное окисление”). В этом цикле АцКоА полностью расщепляется до СО2 и Н2О. При этом образуется 12 АТФ в расчете на молекулу АцКоА, вступившую в цикл. Если рассчитать на 1 молекулу глюкозы, то на 3-м этапе образуется 24 АТФ.
1-й этап проходит 10 промежуточных стадий. В ходе первой части этого этапа молекула глюкозы расщепляется пополам до 2-х молекул фосфоглицеринового альдегида (ФГА).
ОСОБЕННОСТИ ПЕРВОЙ ЧАСТИ 1-ГО ЭТАПА:
Гексокиназа (ГК) работает, чтобы ослабить прочную молекулу глюкозы:


На 3-й стадии фруктозо-6-фосфат еще более ослабляется фосфофруктокиназой (ФФК) и образуется фруктозо-1,6-бисфосфат:

Фосфофруктокиназа — это ключевой фермент ГБФ-пути. Он является «пунктом вторичного контроля». Vmax ФФК больше, чем Vmax ГК. Поэтому, когда глюкозы поступает много, ГК лимитирует скорость всего ГБФ-пути.
Избыток АТФ и избыток цитрата сильно ингибируют ФФК. В этих условиях лимитирующим ферментом ГБФ-пути вместо гексокиназы становится ФФК. Из-за угнетения ФФК накапливаются глюкозо-6-фосфат (Г-6-Ф) и фруктозо-6-фосфат(Ф-6-Ф). Г-6-Ф ингибирует гексокиназу, уменьшая утилизацию глюкозы клеткой и одновременно активирует гликогенсинтетазу.
Если нет избытка АТФ и цитрата, а есть избыток АДФ, то АДФ активирует ФФК, и тогда скорость всего ГДФ-пути лимитируется опять гексокиназой.
В результате фосфофруктокиназной реакции молекула фруктозо-1,6-бисфосфата дестабилизируется (ослабляется) настолько, что сразу распадается на 2 триозы при участии фермента альдолазы (4-я реакция):
Читайте также: Ткань креп шифон стрейч


В следующую (шестую) реакцию ГБФ-пути вступает только ФГА. В результате уменьшается его концентрация и равновесие 5-й реакции сдвигается в сторону образования ФГА. Постепенно весь ФДА переходит в ФГА, и поэтому количество АТФ, синтезировавшееся в последующих реакциях ГБФ-пути, мы учитываем в расчете на 2 молекулы ФГА и других промежуточных метаболитов, которые из него образуются.
В 1-й части 1-ого этапа (от глюкозы до ФГА) расходуется 2 молекулы АТФ: одна — в гексокиназной реакции, другая — в фосфофруктокиназной (3-я реакция первого этапа ГБФ-пути). 2-я часть 1-го этапа начинается с окисления ФГА до ФГК (фосфоглицериновой кислоты) в 6-й реакции.

Эта реакция катализируется ферментом «глицеральдегидфосфатдегидрогеназа». Отщепляемый водород передается на НАД с образованием НАДН2. Энергии, которая выделяется при этом окислении, хватает и на то, чтобы одновременно обеспечить присоединение фосфата к альдегидной группе. Присоединяется фосфат макроэргической связью. В результате образуется 1,3-дифосфоглицериновая кислота (1,3-бисфосфоглицерат).
7-я реакция: субстратное фосфорилирование.

Фосфат с макроэргической связью передается на АДФ с образованием АТФ. В результате 7-й стадии в молекуле фосфоглицериновой кислоты остается 1 остаток фосфорной кислоты.
8-я реакция: Фосфат переносится из 3-го во второе положение и образуется 2-фосфоглицериновая кислота.


От 2-фосфоглицериновой кислоты отнимается Н2О. Это приводит к перераспределению молекулярной энергии. В результате на фосфате во втором положении накапливается энергия и связь становится макроэргической. Получается фосфоенолпируват(ФЕП).
10-я реакция: Субстратное фосфорилирование. Фосфат переносится на АДФ с образованием АТФ. ФЕП переходит в ПВК (пировиноградную кислоту).

На этом 1-й этап ГДФ-пути заканчивается, ПВК уходит в митохондрию и вступает во второй этап ГДФ-пути.
Итоги 1-го этапа: 10 реакций, из которых первая, третья и десятая реакции необратимы. Сначала расходуется 2 АТФ на 1 молекулу глюкозы. Потом окисляется ФГА. Энергия реализуется в ходе 2-х реакций субстратного фосфорилирования: в каждой из них образуется по 2 АТФ. Следовательно, на каждую молекулу глюкозы (на 2 молекулы ФГА) получается 4 АТФ путем субстратного фосфорилирования.
Суммарно все 10 стадий можно описать следующем уравнением:
НАДН2 по системе митохондриального окисления(МтО) передает водород на кислород воздуха с образованием Н2О и 3 АТФ, но 1-й этап протекает в цитоплазме и НАДН2 не может проходить через мембрану митохондрий. Существуют челночные механизмы, обеспечивающие этот переход НАДН2 через митохондриальную мембрану — малат-аспартатный челнок и глицерофосфатный челнок (смотрите лекции «Биологическое окисление».
В расчете на одну молекулу глюкозы образуется 2НАДН2.

В дополнение к 2 АТФ, получаемым на 1-м этапе путем субстратного фосфорилирования, образуется еще 6 АТФ с участием кислорода, итого — 8 молекул АТФ. Столько АТФ образуется в расчете на каждую расщепленную до ПВК молекулу глюкозы в ходе первого этапа ГБФ-пути.
Если эти 8 АТФ добавить к 30 молекулам АТФ, которые образуются на 2-м и 3-м этапах, то суммарный энергетический итог всего ГБФ-пути составит 38 АТФ на каждую молекулу глюкозы, расщепленную до СО2 и Н2О. В этих 38 АТФ заключено 65 процентов энергии, которая выделилась бы при сжигании глюкозы на воздухе. Это доказывает очень высокую эффективность работы ГБФ-пути.
Из 38 АТФ основная их часть образуется на 2-м и 3-м этапах. Каждый из этих этапов абсолютно необратим и требует обязательного участия кислорода, так как окислительные стадии этих этапов сопряжены с митохондриальным окислением (без него невозможны). Весь ГБФ-путь от глюкозы или гликогена до СО2 и Н2О называют: АЭРОБНЫЙ РАСПАД УГЛЕВОДОВ.
Ключевые ферменты первого этапа ГБФ-пути: ГЕКСОКИНАЗА и ФОСФОФРУКТОКИНАЗА.
Еще одно ключевое звено находится в ЦТК (3-й этап ГБФ-пути). Ключевое звено на 3-м этапе необходимо потому, что АцКоА, вступающий в ЦТК, образуется не только из углеводов, но и из жиров и аминокислот. Следовательно, ЦТК — это конечный «котёл» для сжигания ацетильных остатков, образующихся из углеводов, жиров и белков. ЦТК объединяет все метаболиты, образующиеся при распаде углеводов, жиров и белков.
Ключевые ферменты ЦТК: цитратсинтетаза и изоцитратдегидрогеназа. Оба фермента угнетаются избытком АТФ и избытком НАДН2. Изоцитратдегидрогеназа активируется избытком АДФ. АТФ ингибирует эти ферменты по-разному: изоцитратдегидрогеназа ингибируется АТФ намного сильнее, чем цитратсинтаза. Поэтому при избытке АТФ накапливаются промежуточные продукты: цитрат и изоцитрат. В этих условиях цитрат может выходить в цитоплазму по градиенту концентраций.
2-й и 3-й этапы ГБФ-пути протекают в митохондриях, а 1-й — в цитоплазме.
1-й этап отделен от 2-го и 3-го этапов митохондриальной мембраной.
Поэтому 1-й этап может выполнять свои особенные функции. Эти функции
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом