
Посттравматическое восстановление скелетных мышц является актуальной медико-биологической проблемой. Как правило, после глубоких мышечных повреждений полноценного восстановления ткани не происходит. На его месте формируется грубоволокнистый рубец, что приводит к нарушению функционирования органа. Существующие технологии коррекции данных дефектов – мышечная аутопластика, аллопластика, ксенопластика, клеточные технологии, генная терапия являются трудоемкими, травматичными и сопряжены с осложнениями [1]. Одним из перспективных направлений в регенеративной медицине являются тканевая инженерия с использованием биодеградируемых трансплантатов [3]. Биоматериалы Аллоплант в различной модификации зарекомендовали себя как эффективные стимуляторы регенерации соединительной ткани [6]. Целью исследования явилось определение морфологических аспектов регенерации скелетной мышечной ткани после механического повреждения с использованием одного из его видов – аллогенного губчатого биоматериала (АГБ).
Материалы и методы исследования
Для исследования использовали половозрелых крыс породы Вистар. Работу проводили с соблюдением «Правил проведения работ с использованием экспериментальных животных» (приказ Минвуза от 13 ноября 1984 г. № 724). В опытной серии (n=36) после разреза кожных покровов на задней поверхности голени производилось выделение икроножной мышцы и пяточного сухожилия, а также малоберцового нерва, который не повреждали. Затем на брюшко мышцы в средней трети наносили дефект длиной 3–4 мм. В толщу между проксимальной и дистальной культями укладывался АГБ соответствующих размеров и фиксировался нитевидным сухожильным трансплантатом. В контрольной серии (n=36) в области икроножной мышцы был нанесен дефект длиной 3-4 мм. После чего на кожу в обоих случаях накладывали швы Vicryl 6–0. Малоберцовый нерв не повреждали. АГБ был изготовлен в данном случае из сухожилий крысы и обработан методом лиофилизации, что позволяет добиться модификации структуры в губчатую форму с увеличением объема в 6 раз [7]. Нитевидный сухожильный трансплантат, также был аллогенного происхождения, изготовлен из сухожилия крысы и представлял собой нить, которая применялась для фиксации трансплантируемого АГБ к культям мышцы. Из опыта животные выводились путем инсуфляции летальной дозы паров раствора фторотана. Забор биопсийного материала проводили через 3, 7, 14, 30, 60 и 90 суток после эксперимента. Все трансплантаты были обработаны по оригинальной запатентованной технологии АллоплантÒ, разработанной в ФГБУ «Всероссийский центр глазной и пластической хирургии МЗ РФ» (Патент РФ на изобретение № 2189257, ТУ 9398-001-04537642-2011). Гистологические срезы тканей окрашивали гематоксилином и эозином, по Ван Гизону и по Маллори. Микроскопические исследования проводились с использованием светового микроскопа AxioImager Z1, оснащенного фотонасадкой ProgRes C3 и программой анализа изображений Axiovision (C. Zeiss, Германия). Для электронномикроскопического исследования кусочки тканей фиксировали в 2,5 %-м р-ре глютаральдегида, приготовленного на какодилатном буфере (рН 7,2–7,4) с дофиксацией в 1 %-ном р-ре OsO4 на том же буфере. Материал обезвоживали в спиртах возрастающей концентрации и заливали в эпон-812 по общепринятой методике. Предварительно готовили полутонкие срезы на ультратоме ЕМ UС 7 (Leica, Германия) и окрашивали их раствором толуидинового синего на 2,5 %-ном р-ре безводной соды. На данных срезах выбирали участки для электронномикроскопического исследования. Ультратонкие срезы контрастировали 2 %-ным водным р-ром уранилацетата, цитратом свинца по Рейнольдсу и изучали в трансмиссионном микроскопе JEM-1011 (Jeol, Япония) при ускоряющем напряжении 80 кв.
Результаты исследования и их обсуждение
В контрольной группе в начальные сроки в ране обнаруживались очаги острого воспаления и кровоизлияния. В период 7-14 суток отмечалось развитие пролиферативной стадии воспаления. Дефект замещался грануляционной тканью, представленной толстыми фуксинофильными пучками коллагеновых волокон, инфильтрированных соединительнотканными и иммуногенными клетками. Среди клеток соединительной ткани преобладали клетки фибробластического ряда: мезенхимные клетки, фибробласты с активной коллагенсинтетической деятельностью (коллагенобласты II типа). В меньшей степени выявлялись лимфоциты, макрофаги, преимущественно секреторного типа, и их производные – эпителиоидные клетки и гигантские клетки инородных тел. Отмечался скудный васкулярный рисунок. Резецированные мышечные волокна запечатывались коллагеновыми волокнами за счет наплыва сарколеммы. Через 21 сутки в месте дефекта выявлялись признаки трансформации грануляционной ткани в жировую за счет терминальной дифференцировки фибробластов в адипоциты. В цитоплазме фибробластов помимо характерных резко расширенных каналов гранулярного эндоплазматического ретикулюма выявлялись многочисленные разнокалиберные липидные капли (рис. 1).

Рис. 1. Фибробластическая клетка адипоцитарного направления с хорошо развитой сетью ГЭР. Через 21 сутки после нанесения дефекта в скелетной мышечной ткани. Электронограмма. Увеличение х6000
Спустя 30 суток в области дефекта обнаруживался регенерат, состоящий из мышечной, жировой и плотной волокнистой соединительной тканей (рис. 2).
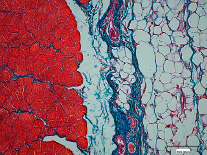
Рис. 2. Жировое перерождение соединительной ткани через 30 суток после нанесения дефекта в скелетной мышечной ткани. Окраска по Маллори
В контрольной группе стадия острого воспаления переходила в фазу пролиферации с образованием гранулематозной ткани, где была наиболее усилена фибробластическая деятельность. Выраженная мезенхимная реакция и дифференциация в коллагенобласты II типа способствовала интенсивному синтезу коллагеновых волокон. Выявлялся их дефицит и дифференциация в неактивные гигантские формы с низкой секреторной активностью. Известно, что фенотипическая незрелость и фагоцитарная инертность макрофагов могла способствовать незавершенному фагоцитозу и привлечению лимфоидных клеток, что обуславливало фиброзирование дефекта в мышечной ткани [2]. Происходила редукция гемокапилляров. Ишемия тканей могла способствовать распаду мышечных волокон и массовой гибели клеток, что также провоцировало развитие фиброза [2]. Результатом заживления мышечного дефекта явилось образование рубца с последующим перерождением в жировую ткань.
В опытной группе через 3 суток после пересадки АГБ в паратравматической зоне обнаруживались дилятация и повышенная проницаемость кровеносных сосудов, нейтрофильная инфильтрация, отек межпучковых пространств, разрушения миоцитов, кровоизлияния. В зоне трансплантата выявлялся фуксинофильный тканевый экссудат, пронизанный фибриновыми нитями в виде тонковолокнистой сети. Признаки острого воспаления были обусловлены механическими воздействиями, возникшими вследствие оперативного вмешательства. Проницаемость кровеносных сосудов способствовала клеточной элиминации, набуханию и ферментативному воздействию на трансплантат. Через 7 суток воспалительная инфильтрация в реактивной зоне и в самом трансплантате менялась на макрофагально-фибробластическую. Выявлялись макрофаги фагоцитарного типа, а фибробласты с умеренно расширенными каналами гранулярного эндоплазматического ретикулюма классифицировались как коллагенобласты I типа – клетки с умеренным синтезом коллагена. Наряду с соединительнотканными клетками обнаруживались малодифференцированные клетки без определенной специализации, а также клетки миогенной дифференциации – миосателлитоциты II типа и миобласты. После лизиса и резорбции балок АГБ макрофагами происходило постепенное разрастание обильно васкуляризированной рыхлой неоформленной соединительной ткани. Спустя 14 суток происходило замещение АГБ от периферии до центра по всей площади. Наблюдались признаки формирования мышечно-соединительнотканного регенерата с преобладанием рыхлой соединительной ткани. Регенерат был представлен тонкими пучками коллагеновых волокон инфильтрированных макрофагами и фибробластами. Причем, макрофаги присутствовали в наибольшем количестве. Строму сопровождали гемокапилляры, свободные миоциты, образующие почки роста и тяжи новообразованных тонких мышечных волокон (рис. 3).
Читайте также: Полость тела заполнена рыхлой тканью паренхимой это признак
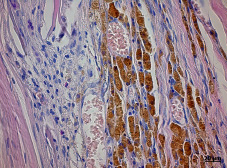
Рис. 3. Массивная инвазия макрофагов в центральной зоне через 14 суток после имплантации в скелетную мышцу крысы губчатого аллотрансплантата. Окраска гематоксилином и эозином
Через 30 суток в очаге трансплантации обнаруживался мышечно-соединительнотканный регенерат с преобладанием мышечной ткани. Пространственная ориентация растущих миосимпластов была параллельна балкам предсуществующего биоматериала. Через 60 – 90 суток регенерат был представлен пучками мышечных волокон окутанных эндо- и перимизием (рис. 4).
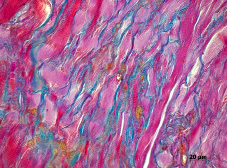
Рис. 4. Новообразованная мышечная ткань в зоне трансплантации губчатого аллотрансплантата спустя 90 суток. Параллельно ориентированные пучки мышечных волокон окутаны эндо- и перимизием. Окраска по Маллори
Биоматериалы Аллоплант изготавливаются из волокнистых соединительнотканных кадаверных тканей. После их имплантации при замещении у реципиента формируется собственная органотипическая рыхлая волокнистая соединительная ткань [5, 6], что подтверждает данное исследование. Между расширенными стромальными элементами свободно мигрировали эндотелиальные клетки гемокапилляров и малодифференцированные миогенные клетки. Продукты резорбции АГБ являются хемоаттрактантами макрофагов и способствуют их фенотипическому созреванию в клетки фагоцитарного типа [5], что наблюдалось в опытной группе. Активированные макрофаги, в свою очередь, влияют на фенотипизм фибробластов, которые ингибируют избыточный синтез коллагена. По данным исследователей, макрофаги также способствуют успешному приживлению миогенных клеток предшественников в раннем периоде заживления скелетной мускулатуры [8, 9, 10]. Происходила ранняя активация миосателлитоцитов и их дифференциация в зрелые миоциты. Новообразованные миосимплаты сопровождали коллагеновые волокна и свободно проникали между ними. За счет удлинения и гипертрофии мышечных волокон, они постепенно вытесняли новообразованную рыхлую соединительную ткань на периферию мышечного пучка. Так формировался эндомизий и перимизий. В данном случае, соединительная ткань выступает «в качестве источника индукционно-формативной тканевой регуляции», а мышечная ткань является регулируемой системой [4].
Таким образом, при использовании АГБ наблюдалось восстановление скелетной мышечной ткани на месте утраченной, в то время как в контрольной группе без применения биоматериала происходило формирование неполноценного соединительно-жировотканного регенерата.
Регенерация различных видов мышечных тканей

Посттравматическая регенерация мышечной ткани является актуальной проблемой в медицине. В течение жизни человек подвергается разным повреждающим факторам, действующим на мышцы, таким как ранения, растяжения, перенапряжение мышц и старческие изменения. Действие этих факторов приводит к различным последствиям: от временного снижения работоспособности мышц до необратимых повреждений и полной потери функции мышцы. Понимание фундаментальных механизмов регенерации мышечной ткани и молекулярных механизмов клеточной дифференцировки в процессе миогенеза позволит управлять регенерацией поперечнополосатых мышц при травме или в процессе возрастных изменений. Кроме этого, понимание отличий механизмов регенерации различных типов поперечнополосатой мускулатуры (скелетной мышцы и кардиомиоцитов), а также роли в регенераторном процессе особого типа клеток – миосателлитоцитов – может помочь в индуцировании реперативных процессов функционально полноценными тканями, в том числе и практически не регенерирующими сократимыми тканями, таких как миокард [1, 2].
Репаративная регенерация развивается после повреждения мышечных волокон. При этом способ регенерации зависит от величины дефекта. В условиях небольшого дефекта мышечного волокна на его концах за счет регенерации внутриклеточных органелл образуются мышечные почки, которые растут навстречу друг другу, а затем сливаются, что приводит к закрытию дефекта. Однако при значительных повреждениях процесс регенерации мышцы приобретает кардинальные отличия. В настоящее время считают, что миосателлиты, впервые описанные Alex Mauro в 1961 году, – главный источник постнатального мышечного роста и репарации мышц [7, 10, 22, 28]. При значительных повреждениях на протяжении мышечного волокна наблюдается усиленная пролиферация миосателлитов в области повреждения и в прилежащих участках, а затем они мигрируют в область дефекта, где формируют миотрубку. Однако репаративная регенерация и восстановление целостности мышечных волокон могут осуществляться только при определенных условиях: во-первых, при сохраненной двигательной иннервации мышечных волокон и, во-вторых, если в область повреждения не попадают фибробласты. Иначе на месте дефекта мышечного волокна развивается соединительнотканный рубец.
На сегодняшний день наибольшее развитие получила теория происхождения миосателлитов у млекопитающих и птиц из параксиальной мезодермы сомитов [5, 26]. Существуют также альтернативные гипотезы происхождения миосателлитоцитов из костного мозга и сосудистых тканей [34]. В пользу происхождения этих клеток из мезодермы сомитов свидетельствуют результаты исследований белков молекулярного каскада, стимулирующего миогенез [37, 43], в которых установлена экспрессия белков семейства MyoD в сомитах и скелетных мышцах во время эмбриогенеза.
Читайте также: Реактивная фолликулярная гиперплазия лимфоидной ткани миндалин
Анатомическое определение миосателлитов изначально опиралось на ультрамикроскопические критерии, и таким образом все клетки, располагавшиеся под базальной мембраной мышечного волокна, считались миосателлитами [24, 25], независимо от функции и экспрессии генов. Недавнее описание молекулярных маркеров различных стадий дифференцировки предшественников миоцитов позволило проводить достоверное определение миогенных клеток методом иммуноморфологии.
В нормальной зрелой мышце сателлитные клетки митотически неактивны и экспрессируют Pax7, c-met, адгезионные молекулы М-кадгерина (Cdh15), сиаломуцин CD34, синдекан 3 (Sdc3) и синдекан 4 (Sdc4), Foxk1, Sox8 и Sox15 [6, 12, 19, 30, 35]. Неактивные сателлитные клетки не экспрессируют миогенные регуляторные факторы, включая белки семейства MyoD.
После активации сателлитные клетки экспрессируют специфичные мышечные факторы транскрипции Myf5 и MyoD с последующей экспрессией миогенина и дифференцировкой [17, 38, 39]. Myf5 может экспрессироваться и в ещё неактивных сателлитоцитах [6]. Фактор Pax7 экспрессируется как в неактивных, так и в пролиферирующих сателлитах [30, 33].
М-кадгерин – кальций-зависимая адгезионная молекула, экспрессируемая в субпопуляции неактивных сателлитных клеток. Предположительные функции этого интегрина заключаются в «заякоривании сателлитов» в их нише и/или миграции сателлитов в место повреждения для регуляции репаративных процессов [6, 19]. Мыши с отключенным геном М-кадгерина имеют нормальное мышечное развитие и регенерацию, что говорит о том, что другие кадгерины, возможно, могут замещать функции М-кадгерина. Это ведёт к предположению об избыточности этих адгезионных факторов. Другими интегринами и адгезионными молекулами, используемыми для определения миосателлитоцитов, являются VCAM-1 и NCAM [9].
Список недавно открытых маркеров, которые можно добавить в список для определения сателлитов, включает в себя лизенин, находящийся в участках сфингомиелина клеточной мембраны, и кавеолин 1 (CAV1) [36].
Возможно, на сегодняшний день, благодаря доступности антител, самым полезным маркером для определения сателлитных клеток является Pax7 [15, 32].
Foxk1 – ядерный транскрипционный фактор, экспрессируемый как в неактивных, так и в делящихся сателлитных клетках, начиная с поздних стадий эмбрионального развития. Было показано, что Foxk1 является вышестоящим регулятором активности ингибитора циклин-зависимой киназы p21, который участвует в регуляции клеточного цикла миосателлитоцитов [13]. Мыши с повреждением гена Foxk1 отстают в росте и имеют дефекты регенерации скелетных мышц, связанные с низкой численностью сателлитов и нарушениями их клеточного цикла [16].
Недавно было высказано предположение, что транскрипционные факторы Sox являются вышестоящими регуляторами Foxk1. Они найдены у всех высших животных и играют ключевую роль в эмбриональном развитии. Из их большого перечня во взрослых мышцах в популяции сателлитных клеток экспрессируются Sox8 и Sox15. Биохимические и эпигенетические исследования позволяют предполагать, что эти факторы ингибируют миогенез через репрессию транскрипции генов семейства MyoD. Мыши, мутантные по гену Sox15, жизнеспособны, но имеют нарушения регенерации мышц [34].
Для выполнения своей роли в поддержании, гипертрофии и восстановлении мышцы миосателлиты должны быть активированы для производства дочерних миобластов. Полученные из сателлитов миобласты характеризуются тем же набором маркеров, что и миобласты, выделенные на любой стадии онтогенеза. После активации миосателлиты быстро начинают экспрессию MyoD, переключаются на другую изоформу CD34, продолжая экспрессировать Pax7, M-кадгерин, и Myf5 [11, 14, 42]. Способные к слиянию миобласты имеют высокий уровень экспрессии десмина, а сгенерированные de novo мышечные волокна и миотрубки экспрессируют эмбриональную изоформу тяжёлой цепи миозина (eMyHC) и продолжают экспрессировать десмин. Появление миогенина означает переход к миогенной дифференцировке и сопровождается появлением разнообразных регуляторных и структурных генов, характерных для скелетных миоцитов [11, 14, 38, 39, 40].
Возрастные изменения значительно влияют на регенераторный ответ. Наиболее распространенным является мнение, что снижение регенеративного потенциала скелетных мышц при старении связано с падением активности миосателлитов. Нарушение их функции как источника миобластов может быть связано с уменьшением клеточного пула сателлитов [24]. Однако, как показали эксперименты, даже сокращенная их популяция способна к эффективному миогенезу in vitro. При этом миосателлиты, ассоциированные со «старыми» мышечными волокнами, были способны к интенсивной регенерации и обеспечению новых сателлитов для пула мышцы-хозяина [22]. Также был показан эффект усиления и «омоложения» сателлитных клеток стволовыми клетками. Первичные миобласты, культивируемые с эмбриональными стволовыми клетками человека (hESC), демонстрируют бурное образование миотрубок. Вокруг колоний клонов hESC формируются исключительно крупные миотрубки, содержащие приблизительно по 50–70 ядер. Образование миотрубок при культивировании с мезенхимальными стволовыми клетками, напротив, не имело особых различий с культивированием изолированных миобластов [34].
Эксперименты показали, что у мышей при старении мышц положительные по Pax7 клетки теряют экспрессию этого гена, при этом направление их дифференцировки изменяется на фибробластический тип, внося таким образом вклад в фиброз [22]. Снижение регенеративного потенциала миосателлитов связывают и с нарушениями сигнального пути Notch за счёт нарушения обмена лиганда Notch-Delta1.
Использование молекулярных маркеров указало на возможную гетерогенность в пуле миосателлитов у молодых мышей [6]. То, что пул миосателлитов может состоять из гетерогенной популяции, было предположено после различных функциональных наблюдений. Во время постнатального роста по темпу деления сателлиты можно разделить на две типичные категории, хотя перечень экспрессируемых ими миогеных маркеров сходен и не позволяет выявить различия. После повреждения мышцы у животного во взрослом возрасте некоторые миобласты экспрессируют миогенин в течение 8 часов и таким образом совершают дифференцировку без или почти без пролиферации, в то время как большинство миобластов практически не делятся раньше 24 часов [27]. Различия в миогенных предшественниках видны также в культуре, где клетки демонстрируют гетерогенность в скорости пролиферации и численности производимых клонов [23]. Наконец, облучение предотвращает рост и поддержание мышц за счёт разрушения большинства сателлитов, но небольшая популяция миогенных предшественников выживает и может снова вступать в восстановительные процессы при повреждении [17, 21].
Читайте также: Размер рулона ткани самый распространенный
В качестве маркера миогенной гетерогенности различных мышечных волокон можно использовать их классификацию по типу тяжелой цепи миозина (MyHC). Например, наличие специфичной для челюстной мышцы кошки сверхбыстрой изоформы MyHC в регенерирующей мышце конечности наблюдается только при пересадке предшественников из челюстной мышцы [18]. Сходным образом у грызунов изоформа MyHC, которую экспрессируют миотрубки, полученные из сателлитов, сходится с изоформой того фенотипа, из которого произошли сателлиты [20, 29]. Данное явление пока не выявлено у людей, и высказано предположение о проявлении этого эффекта только при выращивании культуры на специальных матрицах. Также обращает на себя внимание тот факт, что предрасположенность разных популяций сателлитов к экспрессии различных изоформ MyHC проявляется только после дифференцировки.
Гетерогенность сателлитов в отношении регенераторного потенциала очень заметна для клеток из разных групп мышц. Мышцы головы, например жевательные, регенерируют хуже по сравнению с мышцами конечностей. Это явление может быть связано как с различными сигналами среды в разных мышцах, так и с различиями в самих клетках.
До недавнего времени считалось, что сателлиты унипотентны и что их функция ограничена обеспечением мышцы миобластами для поддержания и репарации. Впоследствии было продемонстрировано, что типично для миосателлитов расположенные в тканях клетки после их изоляции способны дифференцироваться как в миогенные, так и в нейрогенные линии [3]. В связи с этим возник вопрос о правомочности отнесения этих клеток к миосателлитам. Было показано, что сами сателлиты способны дифференцироваться также и в остеогенные и в адипогенные клетки в обычных условиях клеточного культивирования или под действием остеогенных и адипогенных факторов [4, 33].
Для эффективного восстановления структуры и функции мышечной ткани пул сателлитных клеток должен пополняться после каждого акта репарации. Было предложено три сценария, по которым может проходить разделение клеток на две субпопуляции. Изначально предполагалось, что сателлитные клетки гетерогенны и быстро делящиеся клетки участвуют в репарации, в то время как медленные поддерживают пул [27]. Вторая версия предполагала, что сателлитные клетки на самом деле гомогенны и активируются одновременно, но затем принимают разные пути развития, чтобы поддержать и регенерацию мышечных волокон и пула сателлитных клеток. Позже было предположено, что сателлитные клетки могут быть частью иерархической системы и представлять собой миогенные предшественники, ограниченные производством ядер с заменой сателлитных клеток новыми предшественниками, происходящими из стволовых [4, 21, 41].
Для исследования возможности развития этих механизмов в поддержании пула сателлитов были исследованы культивированные мышечные волокна, изолированные с их собственными миосателлитами. При содержании этих волокон в культуре ассоциированные с ними миосателлиты активируются, пролиферируют и дифференцируются, всё еще получая сигналы от мышечного волокна, при этом мышечные волокна изолированы от потенциальных экзогенных источников миогенных клеток, таких как кровеносная система и соединительная ткань [21]. Данные эксперименты наглядно продемонстрировали, что сателлитные клетки изначально гомогенны по признаку активации и экспрессии генов, характерных для этой стадии, но затем принимают разные пути развития. Это подтверждает модель асимметричного деления [41].
Все сателлитные клетки экспрессируют Pax7 и MyoD через несколько часов после активации. После этого появляется расхождение в фенотипе и поведении клеток. Большинство клеток подвергаются быстрой, но ограниченной пролиферации и прекращают экспрессию Pax7 к началу дифференцировки, что согласуется с предыдущими описаниями экспрессии MyoD [8, 14, 38]. Однако не все из этих Pax7+ и MyoD+ клеток следуют программе экспрессии генов до терминальной стадии дифференцировки. Другие поддерживают экспрессию Pax7, но прекращают экспрессию MyoD, в результате чего выходят из дифференцировки. Они становятся неактивными, пополняя пул сателлитных клеток. Эта модель подтверждается наблюдениями за поведением сателлитных клеток во время регенерации in vivo, где большая их часть быстро делится ограниченное количество раз, а затем вступает в дифференцировку, тогда как другие пролиферируют медленно.
Экспрессировавшись единожды, MyoD запускает самоподдерживающийся каскад миогенной детерминации посредством продукции всех факторов MRF. Способность MyoD направлять немышечные клетки в миогенные линии свидетельствует о его серьёзном потенциале для этого каскада.
Таким образом, миосателлитные клетки обладают уникальными способностями к самовоспроизводству и регенерации высокоспециализированной мышечной ткани. Идея, что миосателлиты можно использовать для ремоделирования других мышечных тканей, в частности миокарда, возникла в середине 1990-х годов. Полученные обнадеживающие результаты доклинических исследований транслировались в клинические испытания. Было доказано, что аутотрансплантация миосателлитов в лечении сердечной недостаточности является реальным и относительно безопасным методом (из осложнений – зарегистрированы аритмии). Однако механизм, посредством которого имплантация миосателлитов может улучшить функцию сердца, не ясен. Успешность терапии зависит от ряда факторов, в том числе доставки зону повреждения, долгосрочного выживания, дифференциации в кардиомиоциты и интеграции в новой микросреде [31].
Дальнейшее изучение миосателлитных клеток, а также разработка способов управления их поведением и дифференцировкой является перспективным направлением современной биологической и медицинской науки и может открыть новые горизонты в управлении регенерацией тканей и органов.
Пивоваров Ю.И., д.м.н., профессор, ведущий научный сотрудник, ФГБНУ «Иркутский научный центр хирургии и травматологии» (ФАНО РФ), г. Иркутск;
Сороковиков В.А., д.м.н., профессор, зам. директора по науке, ФГБНУ «Иркутский научный центр хирургии и травматологии» (ФАНО РФ), г. Иркутск.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом