
Регуляция митотической активности в тканях генетический контроль митоза
• Во многих эукариотических клетках основной контрольной точкой служит переход от G2 в М-период цикла
• С переходом G2-M связана активация нескольких протеинкиназ
G2-фаза клеточного цикла связана с подготовкой клетки к митозу. В течение этой фазы большинство клеток растет, и, таким образом, после деления у них сохраняется постоянное ядерно-цитоплазматическое отношение. Во время этой фазы, т. е. до момента вступления клетки в митоз и начала сегрегации хромосом, также узнаются и корректируются ошибки репликации ДНК. Как клетки начинают митоз, когда завершились все процессы? В настоящем разделе мы рассмотрим роль протеинкиназ во вступлении клетки в митоз.
Основной киназой митоза, способствующей переходу G2-M, является комплекс Cdk1-циклин В. По мере накопления митотических циклинов, они связываются с Cdk1. Комплекс накапливается, однако он находится в неактивном состоянии, поскольку белок фосфорилирован при участии представителя семейства киназ Weel. Если в клетках отсутствует активность Weel, то Cdk1 не подавляется. При этом вступающие в митоз клетки достигают небольшой величины, т. е. являются wee. Киназа Mik1 является гомологом Weel и участвует в фосфорилировании и ингибировании активности Cdk1 при пролонгировании S-фазы. Гомолог Weel, Myt1, локализован исключительно в эндоплазматическом ретикулуме (ЭР), в то время как другие представители этого семейства находятся в клеточном ядре. Эти киназы фосфорилируют тирозиновый (Tyr15) или расположенный рядом треони-новый остаток (Thrl4), что приводит к ингибированию активности фермента.
Дефосфорилирование остатков Tyr15 или Thr14 контролирует активацию Cdk1 и вход в митоз. Этот процесс осуществляется Cdc25 фосфатазой. У делящихся дрожжей ген cdc25, в отличие от weel, является необходимым. Если фосфатные группы не удаляются, то митоз не наступает Для гарантии в клетках млекопитающих присутствуют три разных изоформы Cdc25, которые обеспечивают наступление этого ключевого процесса.
В клетках высших эукариот наиболее подробно исследован один из этапов активации Cdk1, протекающий с участием Cdc25. В этих клетках активация Cdc25 происходит с участием Polo-киназ (PLKs), представителей семейства, отличающегося от CDK. После активации Cdc25 с участием PLK и последующего увеличения активности Cdk1 дальнейшая активация Cdc25 происходит под действием самой Cdk1. Такая форма положительной обратной связи при активации Cdk1 приводит к сильному увеличению активности киназы, что приводит клетку к вступлению в митоз. Активность Weel также отрицательно регулируется за счет фосфорилирования Cdk1 с тем чтобы обеспечить резкую активацию Cdk1, необходимую для вступления клетки в митоз. На рисунке ниже схематически представлены процессы активации Cdk1.
У многоклеточных эукариот вхождение клетки в митоз и сам митоз находятся под контролем двух различных CDK-циклиновых комплексов. Например, в клетках млекопитающих Cdk1-циклин А регулирует некоторые процессы в митозе, включающие конденсацию хромосом и их выстраивание у митотического веретена. Варианты циклина В, обладающие разной локализацией и периодически меняющие активность в цикле, вероятно, регулируют другие процессы митоза за счет фосфорилирования различных субстратов. В результате многолетних исследований было найдено много субстратов Cdk1, некоторые из них являются реальными кандидатами на роль эффекторов Cdk1 в митозе.
Хотя Cdk1 считается основным регулятором митоза, другие представители семейства киназ также играют важную роль в различных митотических процессах, и в настоящем разделе мы рассмотрим три группы таких киназ. К их числу относится уже упомянутое выше семейство PLK. Впервые эти киназы были обнаружены в мутантах Drosophila, у которых были нарушены некоторые процессы митоза, а в дальнейшем PLK были найдены в клетках многих эукариот, где они выполняют роль пускового фактора митоза, участвуют в созревании центросом, образовании веретена, сегрегации хромосом и в цитокинезе. В то время как геном Drosophila и дрожжей кодирует только одну киназу, в клетках позвоночных образуется до четырех разных представителей семейства PLK. При этом наиболее функционально близким к ферменту дрожжей и Drosophila является Plk 1. Все ферменты содержат N-концевой киназный домен и С-концевой участок, включающий одну или несколько консервативных последовательностей, которые называются доменами Polo (или Polo boxes). Строение концевых участков и активация PLK показаны на рисунке ниже.
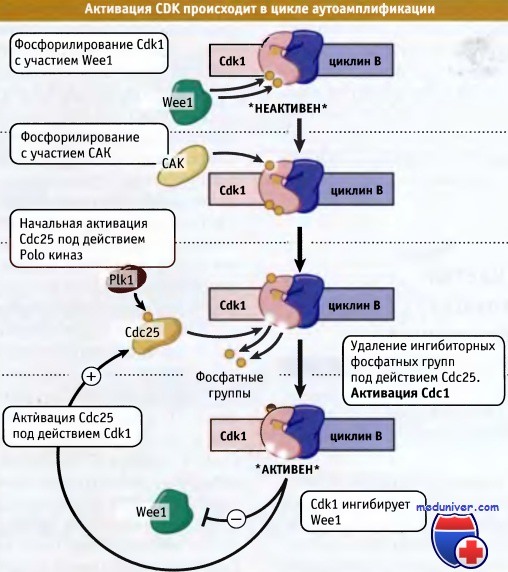
Фосфорилирование Cdk1 подготавливает фермент к активации, однако он поддерживается в неактивном состоянии.
Polo- киназы (Plks) активируют фосфатазу Cdc25, которая затем активирует небольшое количество Cdk1, удаляя фосфатные группы.
После активации эта киназа фосфорилирует Cdc25, что приводит к увеличению уровня активного фермента.
Наряду с этим, при активации некоторого количества Cdc1 она фосфорилирует и инактивирует Weel.
Этот цикл аутоамплификации приводит к резкой активации Cdc1.
Домены Polo выполняют роль внутриклеточных функциональных доменов и участвуют в позиционировании киназ на многочисленных сайтах причаливания, расположенных на центросомах, кинетохорах (белковые структуры, связывающие хромосомы с концами микротрубочек), митотическом веретене и цитокинетическом кольце. Эти домены связываются с мотивом фосфо-Сер/Трео-Про, который представляет собой фосфорилированную консенсусную последовательность в Cdk1 и в других киназах, обладающих сродством к пролин-содержащим сайтам, таких как МАР киназы. Эти данные позволяют предполагать, что PLK присоединяются к белкам, которые предварительно были фосфорилированы Cdk1 или другими киназами, хотя возможно, что in vivo сайты причаливания PLK получают дополнительную информацию от присоединяемых компонентов. Такой механизм мог бы обеспечить координацию активности различных митотических киназ, а также совместную регуляцию Plk1 и других протеинкиназ. Он также помог бы объяснить, почему в митозе PLK активируются параллельно или вслед за Cdkl и другими киназами. По-видимому, в различных сайтах для PLK существует множество субстратов. Все известные субстраты принимают участие в формировании веретена и в цитокинезе.
Читайте также: Ткань бязь для постельного белья размеры
Еще одна группа киназ , участвующих в митозе, представлена семейством NimA-подобных киназ (NEK). Эти ферменты были обнаружены у A. nidulans. История открытия NEK хорошо иллюстрирует тезис о том, насколько важно использовать различные организмы для исследования процессов, контролирующих клеточный цикл. Каждый из объектов исследования обладает своими особенностями, а в совокупности они обеспечивают получение разнообразной ценной информации. Мутанты nimA организма A nidulans заблокированы в С2-периоде, хотя активность Cdk1 в клетках высока. Это говорит о том, что самой по себе активации Cdk1 еще недостаточно для наступления всех событий митоза. Действительно, оказалось, что активность NIMA в митозе меняется параллельно с изменением активности Cdk1. После секвенирования первой обнаруженной киназы гомологичные NEK были найдены в клетках многих эукариот, и было продеменстрировано их участие в нескольких событиях митоза, включая конденсацию хромосом и расхождение центросом.
Третья группа киназ, которая привлекла внимание исследователей в связи с участием в процессах митоза, представлена семейством Aurora. Так же как Cdkl и PLK, киназы Aurora участвуют во многих процессах, включая конденсацию и сегрегацию хромосом, функционирование кинетохора, созревание центросом, образование веретена и цитокинез. Киназы Aurora впервые были обнаружены в почкующихся дрожжах, где они представлены одной киназой. Однако впоследствии было показано их присутствие в клетках многоклеточных эукариот и человека, где они принадлежат к одной из трех следующих групп: Aurora А, В, или С.
Все ферменты семейства Aurora имеют общую структуру: киназному домену предшествует N-концевой домен, а после киназного домена расположен небольшой С-концевой участок. Уровень Aurora А увеличивается на ранней стадии митоза, а при наступлении анафазы он понижается. Снижение содержания фермента, так же как и в случае циклинов, обусловлено его протеолизом с участием убиквитина. Активность Aurora А также регулируется за счет ее фосфорилирования по сайту, расположенному в Т-петле. Как и для Cdk1, это фосфорилирование играет существенную роль в активации Aurora А, и процесс находится под контролем нескольких, пока неполностью охарактеризованных белков. В регуляции активности киназы Aurora А, и, вероятно, Aurora В также участвует протеинфосфатаза 1 (РР1).
Хотя очевидно, что киназы Aurora необходимы для процессов митоза, предстоит еще многое выяснить об их регуляции и о деталях механизма действия. Также неясно, каким образом активность киназ Aurora связана с активностью других киназ, принимающих участие в митозе. Например, неизвестно, функционируют ли они в митозе параллельно, по отдельности или в сочетании с другими киназами.
К числу важнейших выводов, полученных в результате исследований киназ Aurora, относится признание того, что эти ферменты, вероятно, участвуют в генезе опухолей. Опухоли многих типов содержат повышенный уровень киназ Aurora, и гиперэкспрессия Aurora А может приводить к злокачественной трансформации клеток грызунов. Хотя механизм этого явления неизвестен, гиперэкспрессия фермента, вероятно, вызывает амплификацию центросом и дефекты сегрегации хромосом. Более того, считается, что ген, кодирующий Aurora А, обусловливает предрасположенность к развитию рака.
Обобщая все изложенное выше, отметим, что инициация митоза и прохождение клетки через эту стадию цикла требуют функционирования CDK, Plk, NEK и протеинкиназ семейства Aurora. Поскольку активность этих киназ играет критическую роль в поддержании целостности генома, активность каждой киназы регулируется тщательным образом с тем, чтобы избежать несвоевременной активации и обеспечить своевременную ее инактивацию. Накапливаются данные, свидетельствующие о зависимости активности Plk от присутствия других киназ, особенно CDK.
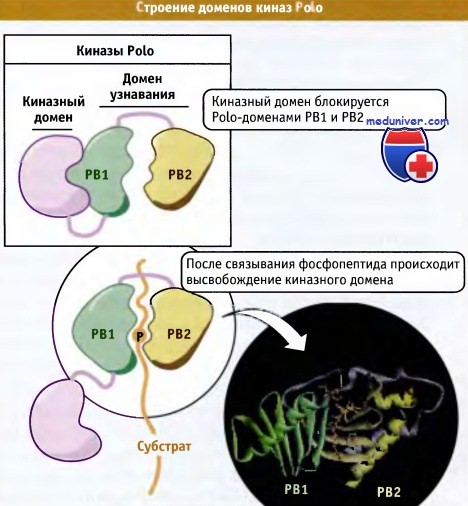
Киназы семейства Polo обладают N-концевым киназным доменом и С-концевым участком,
содержащим два консервативных домена, называемых Polo-домены (РВ1 и РВ2).
Эти домены участвуют в узнавании киназой своих внутриклеточных партнеров.
Читайте также: Камчатка ткань что это такое
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Механизм регуляции митотической активности
Репликация ядерной ДНК происходит в определенный период жизненного цикла клетки – синтетический период интерфазы. Ауторепродукция хромосом и последующее за нее деление клетки направлены но то, чтобы дочерние клетки получили всю полноту генетической информации, имевшуюся в исходной родительской клетке. Чередование двух этих важных процессов составляют основу клеточного цикла. Клеточный цикл означает многократное повторение последовательных событий, занимающих определенный отрезок времени. Обычно циклические процессы изображаются в виде окружности (СЛАЙД № 2).
Фрагмент ДНК от одной точки начала репликации до другой точки образует единицу репликации – репликон. Эукариотические хромосомы содержат большое число репликонов. В связи с этим редупликация хромосом эукариот начинается в нескольких точках. В разных репликонах репликация может идти в разное время или одновременно.
Процесс репликацию ДНК происходит с участием множества ферментов. Слайд № 1. Для осуществления редупликации, цепи материнской молекулы ДНК отдаляются друг от друга. Водородные связи между ними разрушаются геликазой. Область расхождения полинуклиотидных цепей в зонах репликации называют репликационными вилками. В каждой такой области при участии фермента ДНК- полимеразы к освободившимся водородным связям нуклеотидных последовательностей присоединяются свободные нуклеотиды из нуклеоплазмы. Из двух реплицируемых дочерних цепей одна реплицируется непрерывно и синтез ее идет быстрее. Эту цепь назвали лидирующей. Синтез другой цепи идет медленнее, т.к. она собирается из отдельных фрагментов. Такую цепь называют запаздывающей. Ферменты рестриктазы сначала разрезают ее на отдельные фрагменты( фрагменты Оказаки), затем каждый фрагмент реплицируется отдельно и только потом происходит объединение реплицированных фрагментов в единое целое с помощью лигазы.
Ауторепродукция генетического материала у вирусов и бактерий идет на уровне молекул ДНК. У эукариотов материальная основа генетической информации — ДНК, соединяется с белками и образует хроматин. Ауторепродукция хромосом у высших организмов была доказана в опытах Тейлора, Вудса и Хигиса с использованием ауторадиографии хромосом. Опыты этих ученых доказали, что ауторепродукция хромосом высших форм также подчиняется законам ауторепродукции молекул ДНК. Она идет по типу полуконсервативной ауторепродукции. Когда количество ДНК в клетке удваивается, каждая хромосома представлена двумя хроматидами каждая из которых содержит двойную нить ДНК. Ауторепродукция молекулы ДНК составляет основу самоудвоения генетической информации записанной в хромосомах.
Затем клетки переносили на среду, где содержался нормальный изотоп азота (N14). По прошествии времени соответствующей одной генерации кишечной палочки ( примерно 50минут), брали пробы клеток выделяли из них ДНК и центрифугировали. Сначала в ДНК прослеживался и тяжелый азот и нормальный, примерно в равный концентрациях. Через несколько генераций в ДНК обнаруживался только нормальный изотоп азота. Прослеживая такие помеченные полинуклеотидные ДНК, Мезельсон и Сталь показали полную правильность гипотезы Уотсона и Крика.
Годы, последующие за открытием двойной спирали ДНК, ушли на поиски методов, с помощью которых можно было бы доказать существование полуконсервативного способа репликации ДНК, постулированного Уотсоном и Криком. В 1958 году Мезельсон и Сталь проводили опыты на ряде поколений клеток бактерий кишечной палочки. Сначала они выращивали кишечную на среде, содержащей тяжелый изотоп азота (N15) и вся их ДНК метилась этим изотопом.
Согласно модели Уотсона и Крика, предложенной ими в 1953 году, разрыв по водородным связям между комплементарными основаниями ведет к освобождению одноцепочечных полинуклеотидов ДНК. Каждый из них становится матрицей для синтеза новой комплементарной цепи ДНК. Процесс расплетания длинной перевитой молекулы ДНК обеспечить довольно трудно. Для осуществления ауторепродукции необходимы синтетические процессы в цитоплазме, ведущие к образованию четырех типов нуклеотидов, необходимы ферменты для полимеризации полинуклеотидной цепи, необходимы источники энергии и наличие других внутриклеточных условий. Таким образом, ауторепродукция молекул ДНК заключается в широком взаимодействии с белками в зависимости от жизни клетки как целостной системы, и от наличия определенных условий внешней среды.
Молекулы, несущие генетическую информацию, строятся заново в каждом клеточном поколении. В принципах самовоспроизведения молекул ДНК заложена основа устойчивого сохранения всей специфики генетической информации данного вида и данной особи. Это обусловлено комплементарностью при достройке дочерних молекул ДНК, которые имеют идентичные последовательности четырех оснований и, следовательно, то же информационное содержание, что и родительская ДНК.
Клеточный цикл состоит из трех главных стадий: интерфазы, митоза и цитокинеза.
Интерфаза включает G1-период, S-период, G2-период. Промежуток между S-периодом и митозом назвали пресинтетическим (G1— длительностью примерно 4 часа) Далее идет синтетический (S). Он начинается примерно спустя 8 часов после митоза и завершается через 7-8 часов. После синтетического, перед собственно митозом – постсинтетический (G2) происходящим примерно в течение часа.
Читайте также: Маска очки из ткани
Митотическое деление включает 4 фазы: профаза, метафаза, анофаза и телофаза (СЛАЙД № 3).
Самая продолжительная фаза клеточного деления- профаза. На этой стадии интерфазные, деконденсированные и уже редуплицированные хромосомы переходят в компактную форму митотических хромосом- хромосомы (d)
Начинается образование аппарата веретена деления, с участием центриолей и микротрубочек. Веретено деления обеспечивает выстраивание d-хромосом в одной плоскости, в середине клетки и передвижение S-хромосом к полюсам. Ядерная оболочка в конце профазы распадается и цитоплазма и кариоплазма сливаются.
Метафаза. Хромосомы, состоят из из 2-х примыкающих друг к другу S-хромосом (хроматид). Они выстраиваются на экваторе клетке, формируя метафазную пластинку .
Анафаза. Начинается анафаза с расщепления центромерной области каждой из хромосом, в результате чего две S-хромосомы полностью отделяются одна от другой. Благодаря этому каждая дочерняя клетка получает по идентичному набору из 46-ти S-хромосом. После разделения центромер одна из S-хромосом начинает двигаться к одному полюсу, другая к другому.
Телофаза. У каждого полюса происходит диспирализация хромосом в обратном порядке от хромосомного до нуклеосомного уровня. Наконец, наследственное вещество принимает вид глыбок хроматина, либо плотно упакованных, либо деконденсированных.
Вновь формируется ядрышко, ядерная оболочка, окружающая хроматин и кариоплазма.
Цитокинез.В след за делением ядра происходит деление цитоплазмы. В животных клетках происходит образование в кортикальном слое « сократимого кольца» из микрофиламентов. Эти структуры состоят из белков актина и миозина , способных к сокращению. Сокращаясь они обеспечивают разделение цитоплазмы путем инвагинации плазмалеммы.
Таким образом, в результате митотического деления клеток, вновь образованные дочерние клетки идентичные между собой и являются копией материнской клетки, что важно для последующего роста, развития и дифференцировки клеток, тканей.
Поддержание числа клеток на определенном, постоянном уровне обеспечивает в целом гомеостаз.
В организме человека клетки различных тканей и органов обладают разной способностью к пролиферации. Например, клетки красного костного мозга постоянно делятся; клетки большинства внутренних органов во взрослом состоянии не делятся, но способности к делению митозом не утрачивают; нервные клетки у взрослых людей не делятся, а функцию свою повышают за счет увеличения отростков и увеличения соединений между ними) Во многих случаях клеткам подается сигнал о том, что они должны повысить свою функциональную активность, а для этого может понадобиться увеличение числа, клеток. Например, при недостатке в рационе животного Са, клетки паращитовидной железы начинают усиленно делиться, с тем, чтобы увеличение их числа привело к повышению синтеза гормона. Таким образом, понижение той или иной функции может привести к увеличению численности популяции клеток, обеспечивающих эти функции.
Пролиферация клеток регулируется особыми веществами — кейлонами. Кейлоны — это глипопротеиды с мол. массой 30 000 — 50 000. Они обладают специфичностью в отношении клеток разных типов, тканей органов. Считается, что понижение количества кейлонов стимулирует пролиферацию клеток, например, при регенерации.
Амитоз. Прямое деление или амитоз, описано раньше митотического деления, но встречается гораздо реже. Амитоз – это деление клетки, у которой ядро находится в интерфазном состоянии. При этом не происходит конденсации хромосом и образования веретена деления. Формально амитоз должен приводит к появлению двух клеток, однако чаще всего он приводит к разделению ядра и появлению двух- или многоядерных клеток.
Начинается амитотическое деление с фрагментацией ядрышек, вслед за или одновременно делится перетяжкой ядро (или инвагинацией). Может быть множественное деление ядра, как правило, неравной величины (при патологических процессах). Многочисленные наблюдения показывают, что амитоз встречается почти всегда в клетках отживающих, дегенерирующих или не способных дать в дальнейшем полноценные элементы. Так в норме амитотическое деление встречается в зародышевых оболочках животных, в фолликулярных клетках яичника, в гигантских клетках трофобластов. Положительное значение амитоз имеет в процессах регенерации тканей или органов (регенеративный амитоз). Амитоз в стареющих клетках сопровождается нарушениями биосинтетических процессов, включая репликацию, репарацию ДНК, а также транскрипцию и трансляцию. Изменяются физико-химические свойства хроматина ядер клеток, состав цитоплазмы, структура и функции органоидов, что влечет за собой функциональные нарушения на всех последующих уровнях – клеточном, тканевом, органном и организменном. По мере нарастания деструкции и угасания восстановления наступает естественная смерть клетки. Нередко амитоз встречается при воспалительных процессах и злокачественных новообразованиях (индуцированный амитоз.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом