
Термин «мезенхима» (греч. Mesos — средний, enchyma — заполняющая масса) был предложен братьями Гертвигами (1881). Это один из эмбриональных зачатков (по некоторым представлениям — эмбриональная ткань), представляющий собой разрыхленную часть среднего зародышевого листка — мезодермы. Клеточные элементы мезенхимы (точнее, энтомезенхимы) образуются в процессе дифференцировки дерматома, склеротома, висцерального и париетального листков спланхиотома. Кроме того, существует эктомезенхима (нейромезенхима), развивающаяся из ганглиозной пластинки.
Мезенхима состоит из отростчатых клеток, сетевидно соединенных своими отростками. Клетки могут высвобождаться от связей, амебоидно перемещаться и фагоцитировать инородные частицы. Вместе с межклеточной жидкостью клетки мезенхимы составляют внутреннюю среду зародыша. По мере развития зародыша в мезенхиму мигрируют клетки иного происхождения, нежели из перечисленных выше эмбриональных зачатков, например, клетки нейробластического дифферона, мигрирующие миобласты закладки скелетных мышц, пигментоциты и др. Следовательно, с определенной стадии развития зародыша мезенхима представляет собой мозаику клеток, возникших из разных зародышевых листков и эмбриональных зачатков тканей. Однако морфологически все клетки мезенхимы мало чем отличаются друг от друга, и только очень чувствительные методы исследования (иммуноцитохимические, электронно-микроскопические) выявляют в составе мезенхимы клетки различной природы.

Клетки мезенхимы обнаруживают способность к ранней дифференцировке. Например, в стенке желточного мешка 2-недельного эмбриона человека из состава мезенхимы выделяются первичные клетки крови — гемоциты, другие — формируют стенку первичных сосудов, третьи являются источником развития ретикулярной ткани — остова кроветворных органов. В составе провизорных органов мезенхима очень рано претерпевает тканевую специализацию, являясь источником развития соединительных тканей.
Мезенхима существует только в эмбриональном периоде развития человека. После рождения в организме человека сохраняются лишь малодифференцированные (полипотентные) клетки в составе рыхлой волокнистой соединительной ткани (адвентициальные клетки), которые могут дивергентно дифференцироваться в различных направлениях, но в пределах определенной тканевой системы.
Ретикулярная ткань. Одним из производных мезенхимы является ретикулярная ткань, которая в организме человека сохраняет мезенхимоподобное строение. Она входит в состав кроветворных органов (красного костного мозга, селезенки, лимфатических узлов) и состоит из звездчатых ретикулярных клеток, вырабатывающих ретикулярные волокна (разновидность аргирофильных волокон). Ретикулярные клетки неоднородны в функциональном отношении. Одни из них менее дифференцированы и выполняют камбиальную роль. Другие — способны к фагоцитозу и перевариванию продуктов распада тканей. Ретикулярная ткань как остов кроветворных органов принимает участие в кроветворении и иммунологических реакциях, выполняя роль микроокружения для дифференцирующихся клеток крови.
Ретикулярная ткань строма кроветворных органов
С эмбриологической точки зрения кровяная ткань берет свое начало в первичном сосудистом ареале, мезенхимальных островках Вольфа и Пандера, от которых происходят также сердце и сосуды. В следующей главе (г) описаны фазы образования кроветворных органов, соответственно первичная мезобластичсская, затем гепатолиенальная и, на последних месяцах, окончательная костно-мозговая фаза кроветворения. Первичная мезенхимальиая клетка, содержащая информацию о кроветворной пролиферации и дифференциации восстанавливает ряды крови, как материнскими стимулами, так и стимулами, отправляемыми дифференцирующимися плодными тканями.
Привлекает внимание многосторонний потенциал первичной мезенхимальной клетки, которая порождает ретикулярную строму селезенки, узлов, печени и костей.
В послезародышевой жизни мезенхимальная клетка дифференцируется в клетки-штамм, различной направленности в костном мозге и лимфатических органах. Эта ориентация определяется в период зачаточного развития и является результатом продолжительной филогенетической эволюции. На протяжении этого развития костно-мозговая миелоидная ткань находится в зависимости от мезенхимальных, сосудисто-соединительнотканных и костных структур, составляющих частный микроклимат, способствующий дифференцированной пролиферации.
Филогенетически лимфоидная ткань развивается по плану иерархической структурной организации, причем исходно она появляется в центральных, а затем и в периферических лимфоидных органах (Берчану). Центральными органами являются вилочковая железа, у животных, и сумка Фабриция — у птиц, при том эти два органа способствуют «целенаправленности» иммунологически компетентных клеток, носящих характер лимфоцитов Т и Б. План их организации общий и создается вокруг эпителиальных образований эндотелиальной природы, берущих свое начало в пищеварительном тракте. Эти образования составляют эпителиальную ретикулярную строму, переплетающуюся с мезенхимальной стромой, как, впрочем, во всех лимфоидных органах.
Дифференциацию иммуных клеток, равно как и способность реагировать на антигенный стимул определяет их эмбриологическое происхождение, в тесной связи с эпителием, внутридермальной природы (Defendi, Good).
Функциональное различие, требуемое для дифференцированного развития по типу миелоидных или лимфоидных клеток определяется этими структурными взаимосвязями с разновидной стромой: эпителиальным эндотелием, для лимфоидной системы и костносуставными образованиями — для миелоидной. Еще не полностью выявлены взаимоотношения, налаживающиеся при создании этих двух систем. Точно установлена роль вилочковой железы в дифференциации лимфоидной системы, но не выведены размеры ее участия в функциях костного мозга. Однако из патологии известно, что опухоли вилочковой железы определяют медуллярную аплазию, но отсутствует доказательство взаимозависимости функций вилочковой железы и костного мозга при нормальном состоянии.
Читайте также: Узи мягких тканей плечевой области
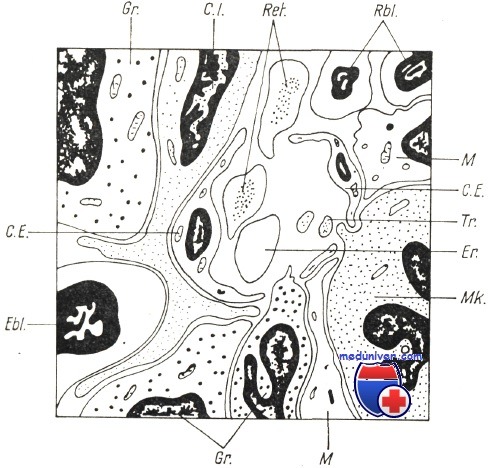
Строение костномозгового синуса.
Ebl—эритробласт; СЕ.— эндотелиальиая клетка; Gr.— гранулоцит; C.I.— интерстициальная клетка; М — макрофаг; Tr — тромбоцит; Mk — мегакариоцит; Еr— эритроцит.
На рисунке изображена функциональная взаимосвязь центральных лимфоидных органов — вилочковой железы и сумки Фабриция — и периферических лимфоидных органов — лимфатических узлов с различной топографической организацией лимфоцитов Т и Б. Лимфоциты Б, а по результатам некоторых исследований и лимфоцаты т находятся и в костном мозге, где образуют мелкие лимфоидные очаги, при этом гуморальный иммунитет находится под контролем лимфоцитов В. Поскольку морфологически клетки-штамм не отличаются от мелких лимфоцитов нам не известно являются ли костномозговые лимфоциты иммунологически компетентными клетками или клетками-штамм.
Проведенными после 1965 г. работами Yoffey и сотр. привели доказательства о том, что лимфоцит это клетка иммуного и в то же время кроветворного восстановления. Методом культур в полунепроницаемых камерах, Григориу и сотр. доказали, что периферические лимфоциты восстанавливают эритробласты. Однако последние данные, полученные в результате анализа колоний в пробирке (Metcalf u Moore) и прижизненно (Мс Colluch) выявили наличие двух различных клеток-штамм, имеющих характеристику лимфоцитов — одну из них в костном мозге, восстанавливающую миелоидные клетки, а другую в лимфоидных органах и периферической крови, восстанавливающую лимфоидные клетки. Эти аргументы выступают в пользу дуалистическоой теории о кроветворении.
Однако не ислючена мысль о способности различной дифференциации той же клетки, в зависимости от структурного, ганглиевого или медуллярного микроклимата, тем самым подтверждая неунистическую гипотезу (Loutit).
В 1979 г. Philips и сотр. впрыскивая клетки-штамм с хромосомными маркерами после облучения проследили возрождение этих клеток у других облученных животных. В связи с этим авторы заключили о наличии полипотентной клетки-штамм (КШп), из которой образуются клетка-штамм для лимфоидных линий Т и В (КШл) и клетка-штамм для миелоидной линии (КШм).
Дифференциацию по той или иной из этих линий определяют условия микроклимата и выделение специфических факторов для организации той или мной из этих линий (Boggs и сотр., Lord и сотр.). При послезародышевой жизни в остальных органах кроветворение не представляется возможным без специфического микроклимата равно как и там, где существуют угнетатели кроветворения. Однако его можно индуцировать при одновременном переносе питательной сети ретикулярной стромы (Humar и сотр.).
Все же функциональные связи и взаимообусловленность возможны в силу собственно структуры костного мозга, по сравнению со структурой ганглиев. В обеих структурах имеется ретикулярная строма с наличием ретикулярных и макрофаговых клеток; существуют также лимфоидные структуры, такие как паренхима ганглиев, но и лимфоидная ткань — в виде мелких фолликулов — и в костном мозге. Нам не известно выполняют ли последние и непосредственную роль в кроветворении посредством определенных общих клеток-штамм или путем стимулирующего трофического влияния на пролиферацию и дифференциацию (Берчану).
Частная патология крови выявит, что заболевания лимфоидного и миелоидного рядов различны. Глубокое поражение стромы и изменение первичной ретикулярной клетки определяет, тем не менее, нарушение обеих систем. Так, при острой недифференцированной лейкемии, рассеянной ретикулосаркоме (PC), нагрузочном ретикулезе, опухолевом гистиоцитозе поражаются обе системы с тяжелыми нарушениями и синдромами недостаточности кроветворения и иммунитета. Структурные связи в костном мозге обусловливают лимфоидные злокачественные метаплазии, смещение миелоидных клеток. При хронической лимфатической, острой лимфобластической лейкемиях или рассеянной лимфосаркоме также наблюдается смещение кроветворной ткани.
Однако существуют патологические аргументы, говорящие о дифференцированной структурной организации этих двух систем. Так, первичная или вторичная аплазия костного мозга не сопровождается аплазией лимфоидной системы; аплазия лимфоидной системы при иммунодефицитных заболеваниях в принципе не поражает кроветворную миелоидную систему. Тем не менее существуют — в настоящее время хорошо известные в иммунопатологии — комплексные иммунодефицитные заболевания, одновременно поражающие обе системы. Так, синдром ретикулярной агенезии и определенные заболевания дисгаммаглобулинемией сопровождаются одновременным нарушением иммуной лимфоидной системы и расстройствами процессов возрождения зернистых и тромбоцитных клеток (Берчану).
Функциональная взаимосвязь в норме, равно как и взаимообусловленность при патологии крови и иммуноклеточной системы говорят в пользу дифференциации миелоидной и лимфоидной систем. Однако эту дифференциацию следует рассматривать, в основном, как результат определенной степени приспособительного отбора по сравнению с условиями среды и структурного микроклимата в послезародышевой жизни. Вместе с тем организационная, генетическая и регенеративная взаимосвязи в зародышевой жизни свидетельствуют о структурном и функциональном единстве организации крови, рассматриваемой как высокодифференцированная мезенхимальная ткань.
Читайте также: Покровная ткань растений где она находится
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
РЕТИКУЛЯРНАЯ ТКАНЬ
Ретикулярная ткань (textus connectivus reticularis (LNH); лат. reticulum сетка; син. сетчатая ткань) — разновидность соединительной ткани, состоящей из ретикулярных клеток и ретикулярных волокон, заключенных в основное межклеточное вещество и образующих рыхлую трехмерную сеть, являющуюся основой кроветворных и лимфоидных органов.
Представления о Ретикулярной ткани формировались в течение многих десятилетий, но лишь в последние 10—15 лет, благодаря использованию достижений цитогенетики, радиобиологии, иммунологии, трансплантологии, были получены принципиально новые данные о ее дифференцировке, структуре и функции.
В эмбриогенезе Ретикулярной ткани дифференцируется из мезенхимы (см.) и в раннем постнатальном периоде постепенно приобретает строение зрелой ткани.

Ретикулярные клетки (cellulae reticulares) относят к клеткам фибробластического типа, наряду с фибробластами, хондробластами и остеобластами, объединяемыми под названием «механоциты». Ретикулярные клетки (рис. 1) имеют уплощенную, веретеновидную или звездчатую форму с гладкой поверхностью; ядро — угловатое или вытянутое. Степень развития органелл, в частности зернистой эндоплазматической сети и комплекса Гольджи (см. Гольджи комплекс), вариабельна и зависит от функционального состояния ретикулярной клетки. Соседние ретикулярные клетки или отростки одних и тех же клеток контактируют друг с другом посредством соединений типа промежуточных или десмосом (см.).
Ввиду того, что ретикулярные клетки часто трудно различить среди массы кроветворных клеток, их идентификация возможна только при использовании электронной микроскопии (см.), гистохимических методов исследования (см.) и методов иммуногистохимии (см. Иммуноморфология) или комбинации этих методов, позволяющих с высокой степенью достоверности выявлять ретикулярные клетки, а также идентифицировать их от внешне сходных с ними клеток системы мононуклеарных фагоцитов (см.).
Гистохимические свойства ретикулярных клеток определяются органными, а также видовыми особенностями. У человека ретикулярные клетки белой пульпы селезенки (см.) отличаются от макрофагов (см.) более низкой активностью эстеразы (см.) и кислой фосфатазы (см.), а от интердигитирующих клеток (разновидности мононуклеарных фагоцитов) — отсутствием активности АТФ-азы. В ретикулярных клетках лимфоцитарной короны лимф, фолликулов селезенки выявляется отчетливая активность 5′-нуклеотидазы (табл.).
Активность 5′-нуклеотидазы определяется также в ретикулярных клетках белой пульпы селезенки крыс и морских свинок, но отсутствует у кроликов. Ретикулярные клетки костного мозга мышей и крыс характеризуются активностью щелочной фосфатазы, локализующейся на их плазматической мембране. Этим признаком они отличаются от макрофагов, в к-рых выявляется активность кислой фосфатазы, локализующейся в лизосомах (см.). Такой морфол. тип ретикулярных клеток часто называют фибробластическим.

В светлых (герминативных) центрах лимфатических фолликулов селезенки и лимфатических узлов (см.) описан особый тип ретикулярных клеток — дендритные ретикулярные клетки. Их отличают крупные размеры, ядро неправильной формы с выраженным ядрышком, многочисленные гладкие пузырьки в цитоплазме. Характерным признаком дендритных ретикулярных клеток являются длинные ветвящиеся отростки с многочисленными складками, глубоко проникающие между окружающими лимфоидными клетками (рис. 2). Отростки дендритных ретикулярных клеток вместе с цитоплазматическими выпячиваниями иммунобластов (В-лимфоцитов) формируют сложную сеть в виде лабиринта. В межклеточном пространстве среди отростков локализуются микровезикулярные структуры и глобулярные плотные частицы диаметром 20—70 нм. В области контакта отростков дендритных ретикулярных клеток видны структуры типа десмосом (см.). В отличие от макрофагов с окрашивающимися включениями и других мононуклеарных фагоцитов дендритные ретикулярные клетки имеют более низкую активность эстеразы и кислой фосфатазы, но высокую активность 5′-нуклеотидазы. По ряду гистохимических признаков они могут быть сходны с мононуклеарными фагоцитами: макрофагами красной пульпы селезенки, металлофильными (аргирофильными) клетками краевой зоны лимф, фолликулов, макрофагами с окрашивающимися включениями. Решающим критерием для идентификации дендритных ретикулярных клеток является способность этих клеток связывать (но не фагоцитировать) иммунные комплексы на своей поверхности.
Ретикулярные волокна (fibrae reticulares), входящие в состав Р. т., состоят из фибрилл различного диаметра, заключенных в гомогенное, плотное основное межклеточное вещество. Фибриллы диаметром 20—50 нм имеют осевую исчерченность, характерную для зрелого коллагена. Тонкие фибриллы диаметром 10 нм локализуются по периферии ретикулярных волокон. Ретикулярные волокна всегда окутаны цитоплазмой ретикулярных клеток в виде чехла (инвагинированы в цитолемму), просвет к-рого всегда сообщается с межклеточным пространством. В лимфоидных органах сеть ретикулярных волокон развита сильнее, чем в костном мозге, причем в тимус-зависимых зонах лимфатических узлов она более рыхлая, чем в корковом и мозговом веществе.
Читайте также: Что такое реобаза ткани
По данным биохимического и иммунохимического анализов, ретикулярные волокна состоят из коллагена III типа (см. Коллаген) и неколлагенового компонента в виде аморфного межфибриллярного вещества с выраженными иммуногенными свойствами. В составе неколлагенового компонента идентифицированы белки (90%), углеводы (4%) и липиды (4%). Для выявления ретикулярных волокон широко используют различные варианты импрегнации серебром. В повседневную практику начинают входить иммуногистохимические и иммунофлюоресцентные методы выявления ретикулярных волокон (см. Иммуноморфология, Иммунофлюоресценция), в основе к-рых лежит использование специфических антисывороток к коллагенам разных типов.
Р. т. образует строму и является носителем специфических органных функций костного мозга и периферических лимфоидных органов (селезенки, лимф, узлов, солитарных и групповых лимф, фолликулов жел.-киш. тракта). После того как было экспериментально доказано, что ретикулярные клетки гистогенетически независимы от кроветворных, их стали относить к категории стромальных элементов, ответственных за создание специфического микроокружения, обеспечивающего миграцию, сортировку, репликацию и дифференцировку кроветворных и лимфоидных клеток. Ретикулярные клетки способны восстанавливать исходное микроокружение при повреждении органов, содержащих Р. т., или переносить его при эктопической трансплантации. Во всех случаях сначала восстанавливается ретикулярная строма, а затем она репопулируется (заселяется) кроветворными или лимфоидными клетками (см. Лимфоидная ткань). В костном мозге ретикулярные клетки формируют также адвентициальный слой синусоидных капилляров и в условиях нормального кроветворения покрывают до 60% их поверхности. Длинные ветвящиеся отростки ретикулярных клеток вступают в специфическое взаимодействие с кроветворными клетками гранулоцитарно-го ряда дифференцировки (см. Кроветворные органы). Концентрация ретикулярных клеток повышена вблизи эндоста.
Экспериментальные данные свидетельствуют о сохранении в пост-натальном онтогенезе гистогенети-ческой близости между разными типами механоцитов (фибробластами, хрящевыми, костными, ретикулярными клетками). Так, при культивировании костного мозга in vitro возникают колонии-клоны фибробластов; нек-рые из этих колоний при обратной трансплантации в организм формируют костномозговой орган (участок костной ткани, окружающей костный мозг), заселяемый кроветворными клетками. Эти данные показывают, что среди ретикулярных клеток костного мозга имеются элементы, способные трансформироваться в истинные фибробласты (судя по их способности синтезировать коллаген I и III типов), и в то же время, проявляющие остеогенные свойства. В другой экспериментальной модели костномозговой орган возникает в результате последовательных морфогенетических процессов под влиянием индуктора — деминерализованного костного матрикса, имплантированного в подкожную соединительную ткань. По мнению Редди, Гея, Гея, Миллера (А. N. Beddi, В. Gay, S. Gay. E. J. Miller, 1977), в этом случае под влиянием индуктора происходит последовательная трансформация фибробластов в хрящевые, костные, а затем ретикулярные (стромальные) клетки. По-видимому, ретикулярные клетки костного мозга способны также трансформироваться в жировые клетки, участвующие в создании кроветворного микроокружения (в красном костном мозге). В норме ретикулярные клетки отличает высокая радиорезистентность, и они практически не делятся.
Существует мнение, что на поверхности дендритных ретикулярных клеток имеются мембранные рецепторы для иммуноглобулинов (см.), с к-рыми связываются иммунные комплексы антигенов с антителами. Согласно другому мнению, антигены (см.) просто медленно фильтруются через лабиринт, создаваемый отростками дендритных ретикулярных клеток. Связывание антигена происходит быстрее у предварительно иммунизированных животных (через 0,5—2 часа), чем у неиммунизированных (через 4—24 часа). Параллельно изменяется и ультраструктура дендритных ретикулярных клеток. В опытах на кроликах показано, что в процессе формирования светлых центров дендритные ретикулярные клетки трансформируются из фибробластических ретикулярных клеток прилежащей лимфоцитарной короны. При этом ретикулярные клетки теряют активность щелочной фосфатазы (и, по-видимому, способность к волокнообразованию).
С возрастом в Ретикулярной ткани увеличивается количество волокнистых структур. Для костного мозга характерно необратимое замещение ретикулярной стромы жировой тканью и прекращение кроветворения. С возрастными и патологическими изменениями Р. т. тесно связано нарушение функции кроветворных и лимфоидных органов. При воздействии высоких доз ионизирующего излучения на организм Р. т. не регенерирует, а замещается фиброзной. Миелофиброз характеризуется интенсивной пролиферацией фиброгенных клеток и последующим массивным отложением коллагена I, III и IV типов. Показано участие ретикулярных клеток в синтезе фибриллярных белков амилоида (см. Амилоидоз). Поражение собственно ретикулярной стромы, сопровождающееся нарушением кроветворения, не всегда можно выявить морфологически. Поэтому важная роль отводится методам клонирования стромальных клеток in vitro, изучению их взаимодействия с кроветворными и лимфоидными клетками в жидкостных и агаровых культурах.
Таблица. Сравнительная характеристика активности ферментов в ретикулярных клетках и мононуклеарных фагоцитах (макрофагах) белой пульпы селезенки человека По данным Мюллер-Хермелинка (H. К. Muller-Hermelink) и др. (1974)
Активность ферментов в различных клетках лимфатического фолликула селезенки
Ретикулярные клетки лимфоцитарной короны
Дендритные ретикулярные клетки светлых центров