
Клетки пучковой зоны секретируют в кровь у здорового человека два основных глюкокортикоида: кортизол и кортикостерон, причем кортизола примерно в 10 раз больше. Секреция глюкокортикоидов регулируется кортикотропином аденогипофиза. Избыток кортизола в крови по механизму обратной связи угнетает секрецию кортиколиберина в гипоталамусе и кортикотропина в гипофизе. Секреция глюкокортикоидов происходит непрерывно с отчетливой суточной ритмикой, повторяющей ритмику секреции кортикотропина: максимальные уровни гормона в крови у человека отмечаются в утренние часы, а минимальные — вечером и ночью (рис. 6.13). Поступающие в кровь гормоны транспортируются к тканям в свободной и связанной (до 95 %) с альфа2-глобулином плазмы (транскортин) формах. Механизм действия глюкокортикоидов на клетки-мишени представлен на рис. 6.14.
Благодаря растворимости в липидах кортизол проникает через мембрану клетки-мишени и взаимодействует с цитоплазматическим рецептором, образуя лиганд-рецепторный комплекс, что обеспечивает транспорт молекулы гормона в ядро, где кортизол связывается с ядерным рецептором, активируя синтез новых белков и ферментов, тем самым обеспечивая метаболические эффекты. Молекула кортизола может образовывать лиганд-рецепторный комплекс и с мембранными рецепторами. Хотя роль этого процесса в реализации эффектов гормона еще изучается, тем не менее известны быстрые негеномные эффекты гормона на возбудимость нервных клеток, связанные с изменением ионного трансмембранного транспорта, обусловливающие изменение поведения.
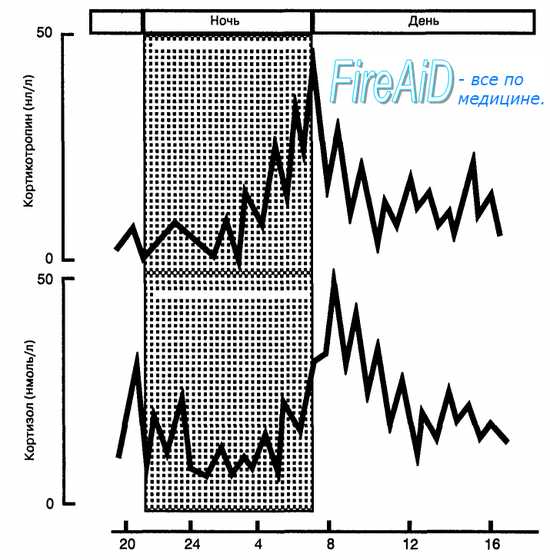
Рис. 6.13. Суточный ритм секреции кортикотропина и кортизола. Максимум концентрации гормонов в крови приходится на утренние часы, при этом прирост уровня кортикотропина опережает рост содержания кортизола.
Среди синтезированных под влиянием кортизола белков в клетке-мишени важнейшую роль для реализации опосредованных эффектов гормона играет семейство липокортинов. Последние, выходя из клетки, связываются со специфическими липокортиновыми рецепторами клеточной мембраны (аутокринный путь влияния), что вызывает подавление активности фосфолипазы-А. Липокортины способны ингибировать фосфолипазу-А и непосредственно, следствием чего является подавление синтеза в клетках простагландинов и лейкотриенов, ослабление их метаболических и регуляторных эффектов. Уменьшение проницаемости клеточных мембран и противовоспалительный эффект кортизола обусловлены ингибированием синтеза лейкотриенов.
Глюкокортикоиды прямо или опосредованно регулируют практически все виды обмена веществ и физиологические функции. Метаболические эффекты глюкокортикоидов проявляются, прежде всего, со стороны углеводного, белкового и жирового обменов. Обобщенно эти сдвиги можно свести к распаду белков и липидов в тканях, после чего метаболиты поступают в печень, где из них синтезируется глюкоза, использующаяся как источник энергии. Эффекты на углеводный обмен в целом противоположны инсулину, поэтому глюкокортикоиды называют контринсулярными гормонами. Гипергликемия под влиянием гормонов возникает за счет усиленного образования глюкозы в печени из аминокислот — глюконеогенеза и подавления утилизации ее тканями. Гипергликемия является причиной активации секреции инсулина. Чувствительность тканей к инсулину глюкокортикоиды снижают, а контр инсулярные метаболические эффекты могут вести к развитию стероидного сахарного диабета.
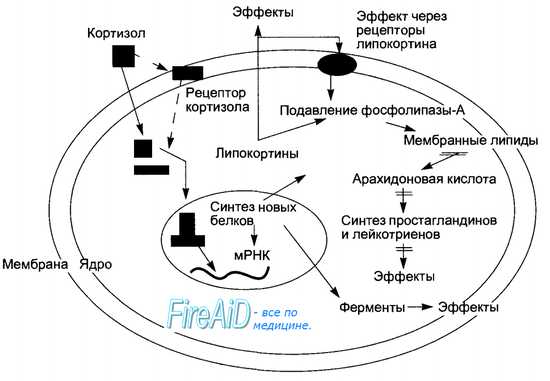
Рис. 6.14. Схема механизма действия кортизола на клетку-мишень. Проникая внутрь клетки через мембрану, молекула гормона последовательно взаимодействет с цитозольным, а затем с ядерным рецептором. Следствие геномного влияния — активация синтеза новых белков, в том числе являющихся внутриклеточными ферментами, что вызывает изменения обмена веществ. К числу синтезируемых под влиянием кортизола белков относятся липокортины. Последние либо выводятся из клетки и взаимодействуют со специфическими для них мембранными рецепторами, либо действуют внутриклеточно. Основной эффект липокортинов — ингибирование мембранного фермента фосфолипазы-А и образования из арахидоновой кислоты простагландинов и лейкотриенов.
На белковый обмен гормоны оказывают катаболический и антианаболический эффекты, приводя к отрицательному азотистому балансу. Распад белка происходит в мышечной, соединительной и костной тканях, снижается уровень альбумина в крови, уменьшается проницаемость клеточных мембран для аминокислот. Однако синтез некоторых белков в печени, например альфа2-глобулинов, глю-кокортикоиды повышают. Со стороны жирового обмена имеют место липо-литический эффект в тканях, гиперлипидемия и гиперхолестеринемия, активация кетогенеза в печени, угнетение липогенеза в печени, стимуляция липогенеза и перераспределения жира в жировой ткани центральной оси туловища и лица, стимуляция аппетита и потребления жира
Читайте также: Евробязь что это за ткань постельное белье
Влияние глюкокортикоидов на реактивность тканей проявляется не только в виде подавления чувствительности к инсулину, но и в повышении чувствительности адренорецепторов к катехоламинам. Глюкокортикоиды вызывают снижение в крови количества лимфоцитов, эозинофилов и базофилов, повышение чувствительности сенсорных структур и возбудимости нервной системы, обеспечение оптимальной симпатической регуляции сердечно-сосудистой системы. Почечные эффекты глюкокортикоидов состоят в стимуляции диуреза путем снижения реабсорбции воды и повышении клубочковой фильтрации; подобно минералокортикоидам, они могут вызывать задержку натрия при потере калия. Глюкокортикоиды повышают синтез ангиотензиногена в печени и тем самым способствуют большему образованию в крови ангиотензина II и секреции альдостерона, увеличивают синтез катехоламинов в мозговом веществе надпочечников. Гормоны повышают устойчивость организма к действию чрезмерных раздражителей, подавляют сосудистую проницаемость и воспаление (поэтому их называют адаптивными и противовоспалительными), из-за катаболизма белка в лимфоидной ткани и угнетения иммунных реакций они оказывают антиаллергические эффекты. Физиологическое влияние кортизола на им-мунокомпетентные клетки носит защитный, регуляторный характер (рис. 6.15).
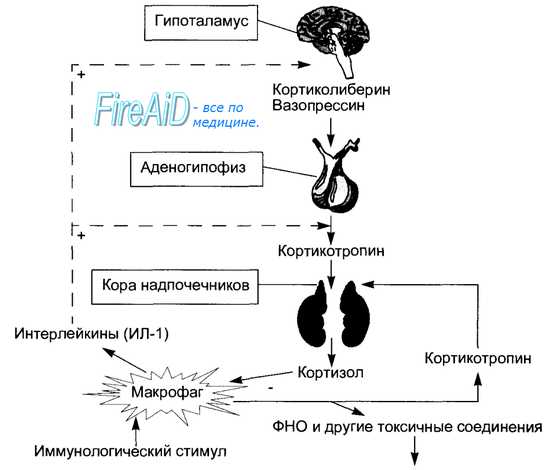
Рис. 6.15. Физиологическое значение влияния кортизола на иммунокомпетентные клетки (на примере макрофага). Кортизол тормозит выделение макрофагом фактора некроза опухолей (ФНО) и других токсичных для клеток организма макрофагальных цитокинов. Под влиянием иммунологического стимула макрофаг выделяет кортикотропин, что усиливает продукцию кортизола надпочечниками и ослабляет выделение макрофагом цитотоксические соединений. Кроме того выделяемые макрофагом интерлейкины, в частности интерлейкин-1, стимулируют гипоталамо-аденогипофизарно-надпочечниковую ось регуляции, что также повышает продукцию кортизола, снижающего цитотоксические эффекты макрофага.
Избыток гормонов в крови вызывает активацию желудочной секреции, выделения НСl, уменьшение числа мукоцитов и продукции слизи, что способствует возникновению язвы желудка — ульцерогенный эффект.
ВЛИЯНИЕ КОРТИЗОЛА НА КОСТИ И МЫШЦЫ
Влияние кортизола на скелетные мышцы
Глюкокортикоиды способствуют атрофии мышц, особенно мышечных волокон типа IIx (которые находятся по своим скоростным свойствам между IIа и IIб) и типа IIб, путем ингибирования синтеза белка и увеличения деградации белка (рис. 8.5; Schakman et al., 2009). Такой синтез ингибируется:
— Не напрямую — комплексом кортизол-глюкокортикоидный рецептор (CGRa) путем подавления активности ферментов, которые отвечают за транспорт аминокислот в мышцы (аминотрансфераз аланина и тирозина).
— Напрямую – кортизол ингибирует синтез белка в мышцах и других тканях несколькими способами:
— Во-первых, кортизол-глюкокортикоидный рецептор (CGRa) блокирует действие транскрипционного фактора переносчика сигнала и активатора транскрипции 5 (STAT5 — Signal Transducer and Activator of Transcription 5) после того, как гормон роста (GH – growth hormone) присоединяется к его рецептору (рис. 8.1): таким образом, кортизол-глюкокортикоидный рецептор (CGRa) может подавлять действие гормона роста (GH) которое (действие ГР) включает в себя экспрессию многих генов белка через транскрипционный фактор (TF) AP-1.
— Во-вторых, кортизол-глюкокортикоидный рецептор (CGRa) блокирует стимулирующее действия инсулина, инсулиноподобного фактора роста I (IGF-I) и аминокислот (особенно лейцина) при трансляции белка путём фосфорилирования белков-репрессоров eIF4E-ВР и серин/треониновой протеинкиназы p70S6K (рис. 2.1, 2.7 и 2.8).

Рисунок 8.5 Геномные действия кортизола приводят к деградации белков. Кортизол, привязанный к GRa прикрепляется к GRE в промоторной области кортизол-чувствительных белков, блокируя их транскрипцию.
Фосфорилирование снижается путем подавления активации Фосфатидилинозит-3-киназы (PI3K)-активации сигнального пути инсулина и инсулиноподобного фактора роста I (IGF-I), серин/треониновой протеинкиназы p70S6K. Это подавляет мишень рампицилина в клетках (mTOR) путем экспрессии регулятора активности генов REDD1 и снижением регулятора активности генов транскрипционного фактора ATF-4, оба которых являются необходимыми для потребления незаменимых аминокислот и синтеза заменимых аминокислоты (рис. 8.5). Подавляя протеинкиназы B (Akt), комплекс кортизол-глюкокортикоидный рецептор (CGRa) также повышает активность киназа-3 гликогенсинтазы (GSK3P), что способствует убиквинтированию транскрипционного активатора P-Катенин и увеличивает экспрессии гена транскрипционных факторов семейства forkhead box класса O (FOXO), который регулируется апоптическими генами atrogin-1, MuRF-1 (Muscle RING-finger protein-1), катепсина L, киназой пируватдегидрогеназы 4 (PDK4) и трансляционного регуляторного белка 4E-BР1 (4E-ВР-1) (Schakman et al., 2009).
— В-третьих, взаимодействие комплекса кортизол-глюкокортикоидный рецептор (CGRa) с транскрипционным фактором NF-kB (NF-kB) приводит к иммуносупрессивному ингибированию синтеза белка. Как показано на рисунке 2.14, транскрипционный фактор NF-kB (NF-kB) активируется, когда воспалительные цитокины вызывают его фосфорилирование IkK киназой. Затем транскрипционный фактор NF-kB (NF-kB) перемещается в ядро, где гликокортикоиды (GRA) блокируют транскрипцию генов провоспалительных цитокинов и их рецепторов (например, фактор некроза опухоли-альфа (TNFa), фактор IL-ip и гранулоцитарный Моноцит колониестимулирующий фактор) путем прикрепления к нему белкового комплекса p65.
Читайте также: Ткань для яхтенных диванов

Рисунок 2.14. Активация транскрипционного фактора NF-кВ. Свободные радикалы и некоторые другие стимулы могут активировать провоспалительные транскрипционные факторы NF-kB через фосфорилирование двух киназ, NIK и IkK. P50/Р65 гетеродимер перемещается в ядро для активации транскрипции генов антиоксидантных ферментов.
Adapted from Free Radical Biology and Medicine, Vol. 28, R.G. Allen and M. Tresini, «Oxidative stress and gene regulation,» pgs. 463-499, copyright 2000, with permission from Elsevier.
— В-четвертых подобно действию на транскрипционный фактор NF-kB (NF-kB), комплекс кортизол-глюкокортикоидный рецептор (CGRa) также ингибирует сигналинг митоза митоген активируемой протеинкиназы (MAPK), содействуя экспрессии гена фосфатазы MAPK.
— В-пятых миогенез также тормозится, когда комплекс CGRa подавляет миогенин и миогенный транскрипционный фактор MyoD, которые участвуют в дифференциации и интеграции сателитных клеток в мышечных волокнах и повышают регуляцию экспрессии отрицательного TF Миостатина (рис. 8.5).
Таким образом, деградация белков инициируется путем активации комплексом CGRa нескольких клеточных протеолитических систем, включая убиквитин протеасомную систему (см. рис. 2.14), катепсин лизосомальную систему, систему каспазы-3 (см. рис. 2.16), а так же СА-кальпин зависимых. Считается, что кальпин отделяет нити актина и миозина, чтобы сделать их доступными для убиквинтирования.
Влияние кортизола на кости
Кортизол может резко снизить плотность костей и увеличить риск перелома кости в случаях, когда имеет место гиперсекреция гормона (например, болезнь Кушинга). Это способствует снижению плотности костной ткани с возрастом, путем преобразования кортизона в кортизол в остеобластах при помощи llpHSDl (Харди & Купер, 2010). Глюкокортикоиды блокируют формирование костной ткани и минерализацию теми же способами, которыми они вызывают атрофию скелетных мышц (Canalis & Delany, 2002). Подавление кортизолом минерализации костей включает в себя связывание глюкокортикоидного рецептора (GRa) с элементом негативной глюкокортикоидной регуляции (nGRE) в промоутере гена остеокальцина (рис. 8.5). GRa, связанная с nGRE в гене-промоутере остеокальцина снижает синтез остеобластов и влияет на связывание рецепторов и связывающих белков (BPs) с анаболическими факторами роста. В частности это действие уменьшает количество остеобластов путем нарушения пролиферации и дифференцировки остеокластов, приводящего к их апоптозу. Два анаболических фактора роста, которые блокируются кортизолом — это инсулиноподобный фактор роста 1 (IGF-I) (рис. 8.5) и трансформирующий фактор роста P (TGFp). Трансформирующий фактор роста P (TGFP) стимулирует синтез белков матрикса коллагена и кости в то время как глюкокортикоиды активируют коллагеназы (MMP ферменты, которые расщепляют коллаген, являющийся основным компонентом костной матрицы). Блокирование кортизолом пролиферации остеобластов, которая в нормальном случае происходит за счет действия IGF, опосредуется транскрипционным фактором C/EBP (C/EBP TF), а также путем супрессии белок-связывающего инсулиноподобног фактора роста 5 (IGFBP-5). Такие факторы роста как: фактор роста фибробластов 2 (FGF-2) и тромбоцитарный фактор роста (PDGF) (которые ускоряют сращение перелома), а так же фактор роста гепатоцитов (HGF) (который способствует регенерации тканей и восстановлению) подавляются кортизолом. Глюкокортикоиды увеличивают остеокластогенез, увеличивая экспрессию лиганд рецептора-активатора ядерного фактора каппа В (RANKL) и снижая экспрессию его рецептора-приманки, остеопротегерина (OPG).
Роль кортизола в метаболизме костной ткани
а) Влияние кортизола на обмен белка. Снижение белков в клетке. Существенным влиянием кортизола на метаболизм белка является уменьшение содержания белков практически во всех клетках организма, кроме клеток печени. Это вызвано как снижением синтеза белков, так и повсеместно возросшим уровнем их катаболизма. Оба эффекта могут быть объяснены снижением транспорта аминокислот в клетке, исключая клетки печени. Возможно, это не главная причина, т.к. кортизол снижает также продукцию РНК и последующий синтез белков, особенно в мышечной и лимфоидной тканях.
Читайте также: Качество ткани с пропиткой
На фоне большого избытка кортизола мышцы становятся настолько слабыми, что больной даже не может встать. Иммунные функции лимфоидной ткани резко падают, составляя ничтожную долю исходных возможностей.
б) Кортизол увеличивает количество белка в плазме крови и в печени. Наряду с повсеместным снижением белков в организме кортизол увеличивает содержание белка в печени. Более того, возрастает уровень белков плазмы крови (продуцируются печенью и высвобождаются в кровь). Это увеличение является единственным исключением на фоне общего истощения резервов белка в организме. Предполагают, что эта разница объясняется возможным влиянием кортизола на поступление аминокислот в гепатоциты (но не в другие клетки организма) и увеличением количества ферментов, необходимых для процессов синтеза.
в) Увеличение содержания аминокислот в крови, снижение поступления аминокислот во внепеченочные клетки и увеличение транспорта в гепатоциты. Исследования, выполненные на изолированной ткани, продемонстрировали, что кортизол понижает транспорт аминокислот в мышечные клетки и, возможно, во все клетки, кроме гепатоцитов.
Снижение поступления аминокислот во внепеченочные клетки снижает в них концентрацию аминокислот с последующим снижением синтеза белков. Кроме того, продолжающийся распад белков высвобождает аминокислоты практически из всех существующих белков. Их диффузия из клеток повышает концентрацию аминокислот в плазме. Кортизол мобилизует аминокислоты из внепепеночных тканей и таким образом истощает резервы белка в тканях.
Увеличение в плазме концентрации аминокислот наряду с возросшим их транспортом в клетки печени под влиянием кортизола дает основание предполагать, что при этом повышается утилизация аминокислот в гепатоцитах, что приводит к увеличению:
(1) скорости дезаминирования аминокислот в печени;
(2) синтеза протеинов в печени;
(3) образования белков плазмы крови в печени;
(4) превращения аминокислот в глюкозу.
Возможно, многие влияния кортизола на метаболические системы организма являются результатом способности кортизола мобилизовывать аминокислоты из периферических тканей с одновременным увеличением ферментов в печени, реализующих ответные реакции печеночных клеток.
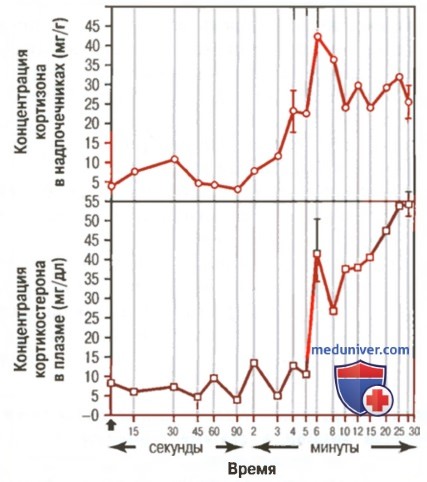
Быстрая реакция коры надпочечников крыс на стресс, вызванный переломом большой и малой берцовых костей в момент, отмеченный 0 (у крысы кортикостерон секретируется вместо кортизола)
г) Влияние кортизола на метаболизм жиров. Мобилизация жирных кислот. Мобилизацию жирных кислот из жировой ткани кортизол осуществляет сходным образом, как и мобилизацию аминокислот из мышц. Увеличение концентрации свободных жирных кислот в плазме повышает их использование для энергетических целей. Возможно, что кортизол оказывает прямое влияние на окисление жирных кислот в клетках.
Механизм мобилизации жирных кислот кортизолом не вполне понятен. Какая-то часть эффектов опосредована снижением поступления глюкозы в жировые клетки. Уже говорилось, что глицерофосфат (один из дериватов глюкозы) необходим для поступления и сохранения триглицеридов в клетках, и его отсутствие в адипоцитах ведет к высвобождению жирных кислот.
Увеличение мобилизации жиров кортизолом наряду с увеличением окисления жирных кислот в клетках обеспечивает сдвиг метаболических систем в клетке в сторону использования жирных кислот вместо глюкозы в качестве источника энергии при голодании и других стрессах. Этот механизм требует для своего формирования несколько часов. Он не является столь же быстрым и настолько же мощным, как эффект похожего сдвига, вызванного снижением инсулина. Тем не менее, увеличение использования жирных кислот в качестве источника энергии является важным моментом в осуществлении долговременных процессов превращения глюкозы и гликогена.
д) Избыток кортизола вызывает ожирение. Несмотря на то, что кортизол способствует умеренно выраженной мобилизации жирных кислот из жировой ткани, у некоторых людей с гиперпродукцией кортизола развивается своеобразный тип ожирения с избыточным отложением жира в шейно-воротниковой зоне, создающим впечатление бычьего торса, и на лице (лунообразное лицо). Хотя причины неизвестны, предполагается, что такой тип ожирения является результатом мощной стимуляции потребления пищи, когда жиры в некоторых тканях образуются быстрее, чем мобилизуются или окисляются.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом