
Жизненный цикл иммунокомпетентных клеток лимфоидной ткани (В- и Т-клетки и их потомки в виде клеток, синтезирующих иммуноглобулины и антигенраспознающие и эффекторные лимфоциты) существенно короче, чем время жизни организма. В связи с этим ведущим условием для осуществления иммунологических реакций служит постоянное протекание в лимфоидной ткани гистогенезов, в ходе которых происходят пролиферация предшественников иммунокомпетеитных клеток и их дифференцировка в зрелые клетки.
Последние, как правило, высокоспециализированы и гетерогенны, а дифференцировочные различия, связанные с иммунологическими функциями, у них жестко закреплены (класс и специфичность синтезируемого иммуноглобулина у В-клеток и специфичность рецепторов для антигенов у Т-клеток). Наоборот, среди клеток-предшественников в течение всей жизни присутствуют и такие, для которых открыты несколько возможностей дифференцировки.
Орган иммунитета — лимфоидная ткань — имеет еще одну важную особенность. Большинство его клеток-предшественников относится к подвижным, рециркулирующим элементам. Они образуются в местах, отдаленных от тех, куда они затем мигрируют и где образуют потомство зрелых клеток.
Таким образом, процессы, которые во многих других тканях осуществляются лишь в момент их закладки при морфогенезе, в лимфоидной ткани протекают всю жизнь. Они обеспечивают постоянное изменение ее клеточного состава, набора синтезируемых иммуноглобулинов (антител) и типов иммунологически активных лимфоцитов в соответствии с меняющимся спектром антигенов, поступающих в лимфоидную ткань.
На лимфоидной ткани удалось убедительно показать, что эти изменения осуществляются главным образом или даже исключительно путем селекции коммитированных клеток-предшественников, предетерминированных к последующей дифференцировке безотносительно к воздействию антигенов. Таким образом, в дифференцировке лимфоидных клеток во взрослом организме естественно выделяются два этапа: развитие антигеннезависимых предшественников и антигензависимая дифференцировка их потомков.
Первый этап начинается с общей стволовой кроветворной клетки и оканчивается коммитированными предшественниками. Неизвестно, однако, в какой последовательности происходят при этом стадии разделения на Т- и В-предшественники, на предшественники, различные по иммунологической специфичности и по классам синтезируемых иммуноглобулинов, а также по принадлежности к разным субпопуляциям Т-клеток. Результаты, полученные при использовании трансплантации костномозговых клеток, маркированных пострадиационными хромосомными перестройками, показали, в частности, что в костном мозге содержатся стволовые клетки, способные к самоподдержанию, но имеющие ограниченные дифференцировочные потенции,— например, такие, которые дифференцируются только в миелоидные или только в Т-клетки.
При этом не обнаружилось пока стволовых клеток, лишенных потенций к миелоидной дифференцировке, но сохранивших свойство быть предшественниками одновременно и для Т- и для В-клеток. Можно поэтому предполагать, что самоподдерживающиеся предшественники Т-клеток (претимические стволовые клетки) прямо происходят из полипотентных стволовых кроветворных клеток, не проходя стадии предшественника, общего для всех лимфоидных клеток. Что касается В-клеток, то между ними и стволовыми кроветворными клетками, возможно, нет вообще предшественников, способных к самоподдержанию.
Вопрос о механизмах, действующих при образовании коммитированных предшественников и регулирующих экспрессию в них генов, ответственных за иммунологические функции, остается открытым. Есть веские основания считать, что этот важный этап развития иммунокомпетентных клеток является антигеннезависимым. Наоборот, последующее развитие уже коммитированных клеток и их потомков отчетливо антигензависимо. Оно обеспечивает изменение клеточного состава лимфоидной ткани путем преимущественного размножения и дифференцировки тех предшественников, чьи поверхностные рецепторы распознают присутствующие в ней в данный момент антигены.
Другой механизм избирательной пролиферации коммитированных предшественников связан с клеточными взаимодействиями, при которых активированная антигеном клетка стимулирует соседние с ней клетки-предшественники. И в том и в другом случае антигены выступают как инициаторы изменения состава клеточных популяций населяющих лимфоидную ткань, и как индукторы синтезируемых этой тканью белков.
Роль лимфоидной ткани в иммунной системе
Непосредственно под мукозным эпителием слизистых оболочек в тесной связи с эпителиальными клетками располагаются лимфоциты пейеровых бляшек кишечника, лимфоидных фолликулов аппендикса, миндалин глотки, лимфоидных фолликулов подслизистого слоя верхних дыхательных путей и бронхов, мочеполового тракта. Все эти лимфоидные скопления получили собирательное название — мукозно-ассоциированная лимфоидная ткань.
В лимфоидных тканях, ассоциированных со слизистыми оболочками, проходят все стадии специфического иммунного ответа в тех случаях, когда антиген проникает в организм через слизистые оболочки. В мукозно-ассоциированных лимфоидных тканях активированные В-лимфоциты дифференцируются в плазматические клетки, продуцирующие специфические антитела, относящиеся к классу иммуноглобулинов A (IgA). Иммуноглобулины А, пройдя через эпителиальные клетки, где они присоединяют секреторный компонент, выходят на поверхность слизистых оболочек в форме секреторного иммуноглобулина A (SIgA), который обеспечивает местную антибактериальную и антивирусную защиту.
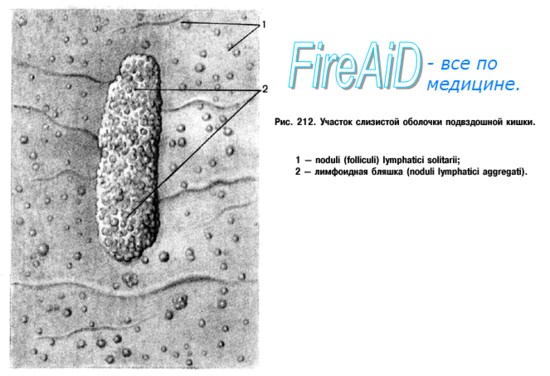
В качестве одного из периферических органов иммунной системы мукозно-ассоциированная лимфоидная ткань является местом:
1) распознавания антигена,
2) антигензависимой пролиферации и дифференцировки Т- и В-лимфоцитов,
3) активации Т- и В-лимфоцитов,
4) продукции цитокинов,
5) продукции и секреции специфических антител — иммуноглобулинов.
Роль лимфоидной ткани в иммунной системе
Концепция общей иммунной системы слизистых оболочек. Под общей иммунной системой слизистых оболочек подразумевают компоненты специфической защиты, реализуемые на внешних слизистых покровах и функционирующие в известной степени независимо от системной иммунологической реактивности. Эта система включает лимфоидную ткань кишечника (GALT-gut associated lymphoid tissue), лимфоидную ткань бронхов (BALT-bronchus associated lymphoid tissue), иммунокомпетентные клетки глотки, слюннных желез, респираторного тракта, молочной железы и гениталий.
Читайте также: Чехлы из ткани для табуреток сшить
Замечательная особенность иммунной системы слизистых оболочек — наличие больших количеств секреторного IgA, что является уникальной характеристикой слизистых покровов. Сывороточные иммуноглобулины и компоненты также присутствуют на различных слизистых покровах и во внешних секретах, но их роль в обеспечении локального защитного механизма менее существенна.
Суть концепции общности иммунной системы слизистых оболочек состоит в том, что выработка секреторных антител происходит не только в местах антигенного воздействия на слизистый покров, но и в отдаленных секреторных участках. Бронхиальный лимфоэпителий, пейеровы бляшки и другие организованные лимфоидные фолликулы изобилуют антиген-реактивными предшественниками В-клеток, коммитированных, главным образом, к синтезу секреторного IgA. Непосредственное антигенное раздражение — необходимое условие для первоначальной активации и пролиферации таких клеток.
Сенсибилизированные антигеном клетки при повторном контакте с тем же антигеном быстро дифференцируются в IgA-иммунобласты, пролиферирующие и мигрирующие сначала в регионарные лимфоузлы, а затем, через грудной лимфатический проток, в кровеносное русло. Во время нахождения в системе кровообращения эти клетки оседают на слизистых оболочках глаз, глотки, гениталий, слюнных и молочных желез, субэпителиальном участке слизистой оболочки верхнего и нижнего отделов респираторного тракта и в lamina propria кишечника.
Специфическая рециркуляция лимфоцитов, стимулированных в лимфоидной ткани слизистых, объясняется тем, что они имеют на своей поверхности молекулы, которые взаимодействуют со специфическими молекулами, размноженными только на поверхности клеток эндотелия венул лимфоидной ткани слизистых, что обеспечивает избирательную рециркуляцию лимфоцитов.
Здесь репопулировавшие клетки начинают активный синтез специфического секреторного IgA, играющего ведущую роль в функционировании общей иммунной системы слизистых оболочек.
Эта концепция имела многочисленные экспериментальные подтверждения. Было установлено, что клетки пейеровых бляшек кролика при введении их облученным аллогенным реципиентам избирательно распределялись в lamina propria и продуцировали антитела преимущественно класса IgA, тогда как клетки, полученные из периферических лимфоузлов доноров, осуществляли специфический homing в периферических лимфоузлах реципиентов и продуцировали антитела преимущественно класса IgG. Сходные исследования выявили, что homing бронхиальных лимфоцитов у реципиентов происходил в BALT.
Оральная иммунизация защищала лабораторных животных от интраназального заражения PC-вирусом. Интраназальное введение дендритных клеток лошадей, праймированных инактивированным герпесвирусом, приводило к появлению иммунитета у мышей.
Иммунизация мышей аналогичным образом инактивированной вакциной против гриппа сообщала устойчивость к интраназальному заражению. Двукратное введение в желудок мышей цельновирионной инактивированной сорбированной вакцины сопровождалось образованием специфических IgA и IgG в легких. При парентеральном введении вакцины в смывах легких IgA не обнаружены.
Энтеральная иммунизация против респираторно-синцитиальной инфекции индуцировала антитела в респираторном тракте, но полную защиту его обеспечивала интраназальная иммунизация. Однако, несмотря на иммунологическую реакцию отдаленных слизистых, наиболее выраженный иммунный ответ образуется в том месте, где слизистая оболочка непосредственно контактирует с вирусным антигеном. Важность локального антигенного стимулирования была убедительно продемонстрирована в экспериментах с вирусом полиомиелита. Уровень специфических секреторных IgA был максимальным в тех участках кишечника, где происходил непосредственный контакт вирусного антигена со слизистой оболочкой.
Живую вакцину Роковак против рота- и коронавирусных энтеритов новорожденным телятам вводили орально перед приемом молозива. После вакцинации заболеваемость и гибель телят от вирусных энтеритов сократились примерно в 4 раза.
В ряде опытов у животных, вакцинированных интраназально, наблюдали образование местных IgA-антител не только в носоглотке, но также в кишечнике и во влагалище. Однако при перректальной вакцинации в кишечнике появлялись IgA- и IgG-антитела в высоких титрах, но не обнаруживались во влагалище. Эти данные указывают, что для индукции местных IgA-антител во влагалище или кишечнике необходима вакцинация именно через эти органы.
Несмотря на то, что исследования по иммунитету слизистых покровов были сосредоточены в основном на изучении секреторных иммуноглобулинов и их роли в функционировании указанной системы, в ряде исследовательских работ указано на участие факторов клеточного иммунитета и, в первую очередь, Т-лимфоцитов в локальном иммунном ответе. Так, установлена миграция и homing Т-лимфоцитов после их антигенной сенсибилизации, а также показана роль Т-хелперов в регуляции местного IgA-ответа.
Представляют интерес данные интравагинальной иммунизации мышей живой вакциной против герпеса. Развивающийся при этом иммунитет может быть перенесен сингенным животным лимфоцитами генитальных лимфоузлов, но не лимфоцитами других лимфоидных органов иммунизированных доноров. Трансплантированные лимфоциты мигрировали преимущественно в ткани половых органов реципиентов, что свидетельствует о важной роли в местном иммунитете вирусиндуцированных Тц-клеток.
Однако имеются наблюдения, когда не были отмечены различия в защите животных от инфекции несмотря на наличие или отсутствие местных IgA-антител после вакцинации.
ЛИМФОИДНАЯ ТКАНЬ
Лимфоидная ткань (лат. lympha чистая вода, влага + греч. eidos вид; син. лимфатическая ткань) — морфофункциональный комплекс лимфоцитов и макрофагов, располагающийся в клеточно-волокнистой соединительнотканной основе и составляющий функционирующую паренхиму лимфоидных органов; Лимфоидная ткань неразрывно связана с гемопоэтической тканью.
К лимфоидным органам, являющимся органами иммуногенеза, относят вилочковую железу (см.), лимфатические узлы (см.), селезенку (см.), лимфоидные элементы костного мозга (см.) и скопления лимфоидной ткани по ходу желудочно-кишечного тракта. Лимфоидные органы в отношении функции иммуногенеза разделяют на первичные и вторичные. Так, вилочковую железу относят к центральным органам системы иммуногенеза (формирование клеточной системы иммунитета), а белую пульпу селезенки, лимфатические узлы и скопления Лимфоидной ткани в слизистой оболочке пищеварительного тракта — к вторичным, или периферическим, органам иммуногенеза.
Читайте также: Основные физиологические функции тканей
Основные этапы эволюции Л. т. достаточно очерчены. У беспозвоночных Л. т. отсутствует. У круглоротых (миног) впервые обнаруживается предшественник вилочковой железы, имеющий вид небольших лимфоидных скоплений в эпителии окологлоточной бороздки. У этих животных установлена способность к иммунному ответу по типу реакции гиперчувствительности замедленного типа и реакции отторжения аллотрансплантата. У примитивных хрящевых рыб обнаруживается селезенка, Л. т. в кишечнике, почках, половых железах, в периваскулярной соединительной ткани. У осетровых рыб появляются плазматические клетки, т. е. способность к выработке специфических антител. Амфибии и рептилии отличаются появлением красного костного мозга, очагов лимфоидного кроветворения в печени. У птиц одновременно с концентрацией лимфоидных элементов в области клоакального органа — фабрициевой сумки — возникают зачаточные лимф, узлы (см. Лимфатическая система).
Все лимфоидные органы млекопитающих, в т. ч. и человека, состоят из соединительнотканной основы, представленной коллагеновыми и аргирофильными волокнами, в к-рых располагаются постоянные (стабильные) клеточные элементы Л. т. и непрерывно мигрируют другие клетки. Основными клеточными элементами Л. т. являются лимфоциты (см.), макрофаги (см.), плазматические клетки (см.), фибробласты, эндотелиальные и ретикулярные клетки. Лимфоциты и их производные, входящие в так наз. лимфоидную систему, и система макрофагов, лежащие в фиброретикулярной ткани, составляют основу Л. т. (так наз. макрофагально-фагоцитарная система). В 1970 г. Ван-Фюрт (R. van Furth) и соавт, предложили данную функциональную систему назвать лимфоретикулогистиоцитарной системой.
Ретикулярные клетки, образующие ретикулярные волокна, по морфологии (на уровне световой микроскопии) не отличаются от фибробластов соединительной ткани (см.). Совр, исследования по гистогенезу кроветворной ткани привели к необходимости пересмотра представлений о ретикулярной клетке. Различные авторы рассматривают ее по-разному. А. Я. Фриденштейн и К. С. Лалыкина (1973) полагают, что ретикулярные клетки стромы вторичных лимфоидных органов (механоциты) образуются из специальных стволовых клеток, отличающихся от стволовых клеток крови. Ретикулярные клетки разных органов, несмотря на морфол, сходство, различаются по направленности дифференцировки: ретикулярные клетки из культур костного мозга при обратной пересадке в организм образуют кость, а из культур селезенки— ретикулярные волокна. И. Н. Кокорин (1970) и соавт, считают, что ретикулярные клетки селезенки являются полипотентными клетками стромы этого органа. Можно предположить, что ретикулярные клетки объединяют несколько типов клеток: гистиоциты, соответствующие макрофагам, моноциты, фибробласты, а также так наз. дендритические и интердигитирующие клетки.
Еще в 1927 г. А. А. Максимов, критикуя концепцию ретикулоэндотелиальной системы (см.), указывал на различия в происхождении, морфологии и функции эндотелиальных клеток сосудов и ретикулярных клеток. Затем было установлено наличие разных предшественников этих клеток и подтверждены их функц, различия. Показано, что эндотелиоциты сосудов, фибробласты и ретикулярные клетки относятся к слабофагоцитирующим элементам: окрашивание их при введении витального красителя (особенно в больших дозах) происходит в основном в результате пиноцитоза (см.), а не фагоцитоза. Оказалось, что в клиренсе крови участвуют преимущественно макрофаги печени (купферовские клетки) и макрофаги красной пульпы селезенки, но не эндотелиоциты и не ретикулярные клетки.
Для обозначения морфофункциональной системы различных клеток лимфоидного и плазматического рядов, участвующих в процессах иммуногенеза, ряд авторов ранее применяли термин «лимфоидно-макрофагальная система». Этим термином объединялись в единую функциональную систему лимфоциты, моноциты, так наз. полибласты, макрофаги воспалительного экссудата, а также плазматические клетки — непосредственные продуценты антител.
Устойчивый уровень процесса физиологической регенерации в первичных лимфоидных органах человека устанавливается в конце эмбрионального периода, во вторичных — в раннем постнатальном периоде.
В процессе функционирования происходит распад и гибель лимфоидных клеток и элементов стромы, число их непрерывно возмещается пролиферацией, дифференцировкой и миграцией клеток.
Наиболее демонстративны возрастные изменения в вилочковой железе, максимальная масса к-рой (30—40 г) приходится на пубертатный период, затем она быстро подвергается инволюции; отмечается нарастание числа лаброцитов (тучных клеток), количество плазматических клеток сначала увеличивается, затем они исчезают, уменьшается количество эпителиоцитов; возникает жировое замещение паренхимы. Нередко после 30 лет лишь микроскопическое исследование позволяет обнаружить в области вилочковой железы небольшие скопления лимфоцитов и эпителиоцитов, заложенных в жировую или фиброзную основу.
Изменения в лимфоидных органах у пожилых людей служат отражением сниженной иммунной реактивности.
Факторы, регулирующие процессы физиол, и репаративной регенерации в первичных и вторичных лимфоидных органах, выяснены частично: соматотропный гормон, кальцитонин, вазопрессин и другие гормоны усиливают пролиферацию клеток первичных лимфоидных органов, гормоны коркового вещества надпочечников подавляют лимфоцитопоэз в селезенке и лимф, узлах; в эксперименте пассивное введение антител тормозит пролиферацию клеток при действии антигена. Установлен гуморальный фактор (тимозин), вырабатываемый вилочковой железой, способствующий пролиферации тимоцитов и образованию О-антигенов на поверхности стволовых клеток костного мозга, мигрирующих в вилочковую железу. Однако окончательно не выявлен фактор, влияющий на лимфоцитопоэз (см. Лейкопоэтины). Не ясны факторы, обусловливающие рост вилочковой железы в постнатальном периоде и инволюцию ее у взрослых организмов. Вместе с тем установлено, что для созревания тимоцитов необходим прямой контакт с чрезвычайно своеобразной ретикулоэпителиальной стромой вилочковой железы. А. Я. Фриденштейн (1973) высказал предположение, что различия в ответе на повреждающее воздействие первичных и вторичных лимфоидных органов могут быть связаны с разницей в характере и происхождении их стромы. Т. А. Рожнова (1971), Борам (К. Borum, 1969) экспериментально установили, что репаративная регенерация в вилочковой железе происходит лишь в тех случаях, когда повреждающее воздействие (облучение, кортизонотерапия, резекция и др.) существенно не нарушает целости стромы. При этом в вилочковую железу мигрируют лимфоидные клетки костного мозга, за счет пролиферации к-рых и происходит регенерация органа, однако его масса уже не достигает нормальной величины.
Читайте также: Узи мягких тканей брюшной стенки что это такое
Репаративная регенерация вторичных лимфоидных органов (селезенки и лимф, узлов) обусловлена не только пролиферацией репопулирующих клеток из костного мозга и вилочковой железы, но и размножением сохранившихся клеток органов. Г. В. Харлова (1975) допускает, что скорость и полнота регенерации вторичного лимфоидного органа зависят от соотношения и созревания в нем Т- и В-лимфоцитов. Показано, что лимфоциты принимают участие в регенеративных процессах не только в лимфоидных органах, но и в печени, легких, коже, почках.
Функциональное значение Лимфоидной ткани определяется ее важнейшей ролью в иммунных реакциях. Процесс выработки антител осуществляется клетками Л. т. Высказывается вполне обоснованное предположение, что Т- и В-лимфоциты путем обратной связи могут контролировать иммунный ответ и деление стволовых клеток. Нек-рые исследователи полагают, что утрата способности Т-лимфоцитов к такому контролю может явиться причиной аутоиммунных заболеваний, а также является одним из возможных условий развития злокачественного новообразования.
Защитная реакция организма на воздействие чужеродных веществ (экзо- и эндогенных) в значительной мере определяется функц, состоянием Л. т., с к-рой связана специфическая (иммунологическая) резистентность (см. Иммунитет). Появляются доказательства большого значения Л. т. не только в иммунных реакциях, но и в неспецифической резистентности организма (см.). Как показали исследования П. Д. Горизонтова (1976), повышение резистентности организма при стрессе характеризуется усилением костномозгового кроветворения, увеличением миграции клеток Л. т. в костный мозг в первые часы после воздействия чрезвычайного раздражителя — так наз. лимфоидный пик. Появление при этом Т-лимфоцитов в костном мозге рассматривается исследователями как свидетельство роли вилочковой железы в неспецифической резистентности организма.
Предполагается, что при неблагоприятных воздействиях распад клеток Л. т. обеспечивает трофическую функцию, т. к. при этом продукты обмена (в частности, нуклеопротеиды) реутилизируются в зонах повреждения тканей, чем компенсируется неблагоприятное воздействие. Бернс (D. W. Bernes, 1962) и соавт, высказали мнение о том, что недостаточность Л. т. и уменьшение ее трофической функции играют важную патогенетическую роль при так наз. болезнях истощения, в т. ч. и при раневом истощении, описанном И. В. Давыдовским.
Заболевания, связанные с поражением Л. т., обычно диагностируются по данным биопсий лимф, узлов (за исключением лейкозов, дисгаммаглобулинемий и аутоиммунных процессов). К заболеваниям Л. т. относятся гипоплазия лимфатических узлов— редкое состояние, отражающее иммунную недостаточность; реактивная гиперплазия лимф, узлов, возникающая при воспалительных процессах и активации иммунных реакций как первичного, так и вторичного характера (см. Иммуноморфология, Иммунопатология). Решение вопроса о характере так наз. воспалительного компонента (появление эозинофилов, нейтрофилов, плазматических и эпителиоидных клеток) нередко представляет значительные трудности.
Особую группу патологических процессов Лимфоидной ткани составляют злокачественные новообразования (см. Гемобластозы, Гистиоцитозы, Лимфогранулематоз, Лимфома, Миеломная болезнь).
Библиогр.: Горизонтов П. Д. Лимфоидная ткань и неспецифическая резистентность организма, Арх. патол., т. 38, № 3, с. 3, 1976, библиогр.; Пестова И. М. Краткий очерк эволюции лимфоидной ткани и её иммуноклеточной реактивности у позвоночных, Арх. анат., гистол, и эмбриол., т. 70, № 3, с. 26, 1976, библиогр.; Структура и функция лимфоидной ткани в онто- и филогенезе, под ред. Е. А. Вагнера и др., Пермь, 1976; Харлова Г. В. Регенерация лимфоидных органов у млекопитающих, М., 1975, библиогр.; Carr J. The line structure of the mammalian lymphoreticular system, Int. Rev. Cytol., y. 27, p. 283, 1970, bibliogr.; он же, The macrophage — a review of ultrastructure and function, L.— N. Y., 1973; Goldstein A. L., Slater F. D. a. White A. Preparation, assay and partial purification of thymic lymphocytopoietic factor (thymosin), Proc, nat. Acad. Sci. (Wash.), v. 56, p. 1010, 1966; Kyriazis A. A. a. Esterly J. R. Fetal and neonatal development of lymphoid tissues, Arch. Path., v. 91, p. 444, 1971; Maximow A. Bindegewebe und blutbildene Gewebe, Handb, mikr. Anat. Menschen, hrsg. v. W. Mollendorff, Bd 2, T. 1, S. 232, B. 1927; Mononuclear phagocytes m immunity, infection and pathology, ed. by R. van Furth, Oxford, 1975.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом