

Учебник описывает последовательность и механизмы реакций основных метаболитических путей энергетического и пластического обмена. Описываются ингибиторы реакций и механизмы их действия, регуляция скорости метаболитических путей. В разработке использовались как классические данные о метаболитических путях, так и новая информация об механизмах реакций и их регуляции. Учебник предназначен для студентов любых ВУЗов для изучения биохимии, в качестве как основной, так и дополнительной литературы.
Оглавление
Приведённый ознакомительный фрагмент книги Биохимия метаболизма. Учебное пособие предоставлен нашим книжным партнёром — компанией ЛитРес.
Пентозофосфатный путь является альтернативным путем окисления глюкозы. Он включает несколько этапов, в результате функционирования которых из трех молекул глюкоза-6-фосфата образуются три молекулы СО2 и три молекулы пентоз. Последние используются для регенерации двух молекул глюкозо-6-фосфата и одной молекулы глицеральдегид-3-фосфата. Поскольку из двух молекул глицеральдегид-3-фосфата можно регенерировать молекулу глюкоза-б-фосфата, глюкоза может быть полностью окислена при превращении по пентозофосфатному пути.
У растений часть реакций пентозофосфатного пути участвует также в образовании гексоз из СО2 при фотосинтезе. Пентозофосфатный путь называют иногда пентозным шунтом, гексозомонофосфатным путем или фосфоглюконатным окислительным путем. Открытие Отто Варбургом (Otto Warburg) в 1931 г. глюкозо-6-фосфат-дегидрогеназы, первого фермента этого пути, сделало возможной его полную расшифровку, которую осуществили Фриц Липман, Фрэнк Дикенс, Бернард Хорекер и Эфроим Рэкер.
Пентозофосфатный цикл не приводит к синтезу АТР, он выполняет две главные функции: 1) образование NADPH для восстановительных синтезов, таких, как синтез жирных кислот и стероидов; 2) обеспечение рибозой синтеза нуклеотидов и нуклеиновых кислот. Ферменты пентозофосфатного пути локализованы во внемитохондриальном пространстве клеткив цитозоле. Как и в процессе гликолиза, окисление осуществляется путем дегидрогенирования, однако акцептором водорода в этом случае служит не NAD, а NADP.

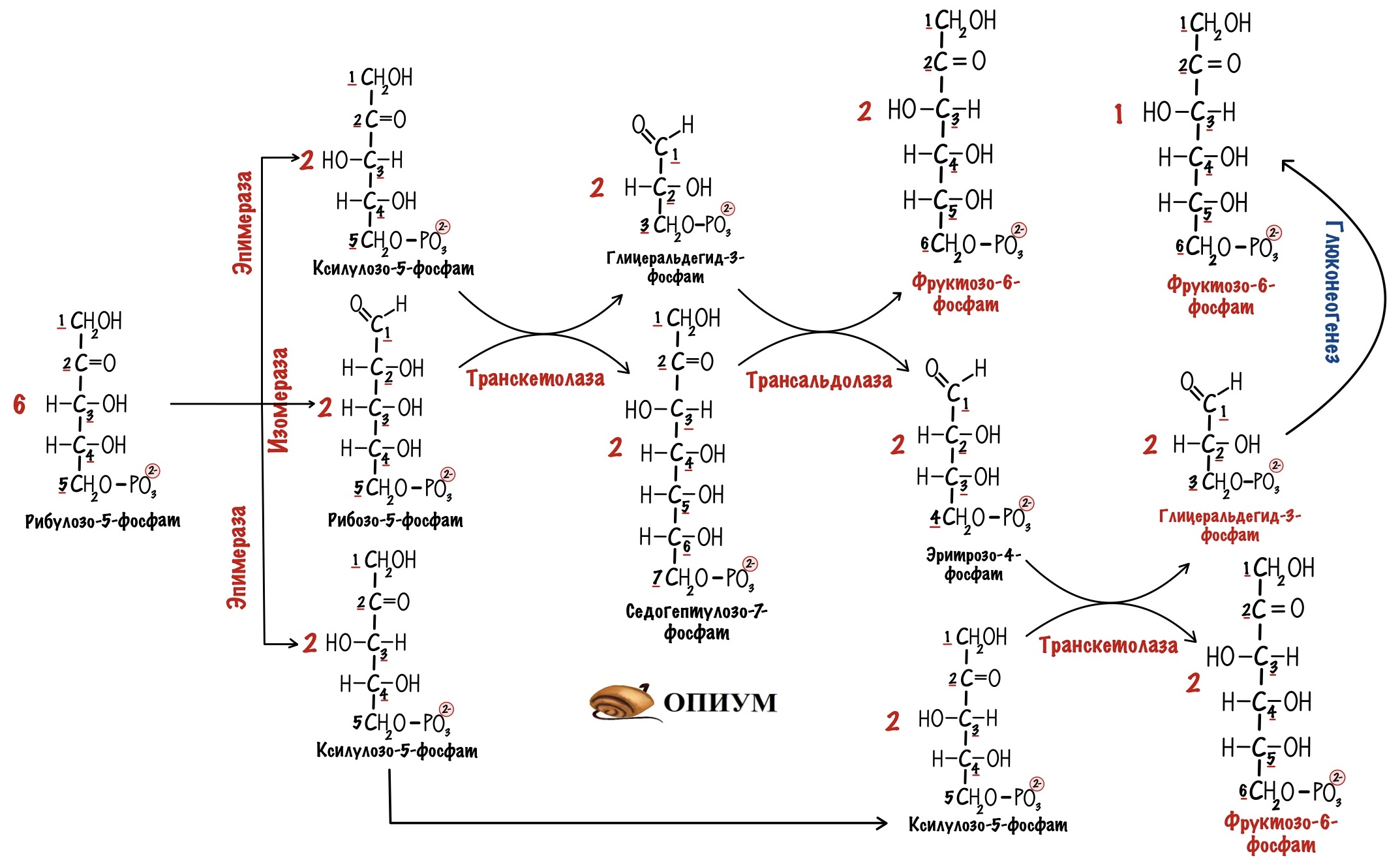
Рисунок 4: Схема реакций пентозофосфатного шунта (черный круг с буквой Р в центре обозначает фосфатную группу).
Последовательность реакций пути можно разделить на две фазы: окислительную и неокислительную (схема реакций представлена на рисунке 4). В реакциях первой фазы глюкоза-6-фосфат дегидрогенируется и декарбоксилируется с образованием рибулозо-5-фосфата. В ходе второй фазы рибулозо-5-фосфат превращается снова в глюкозо-6-фосфат в результате серии реакций, в которых главную роль играют два фермента: транскетолаза и трансальдолаза
Окислительная фаза пентозофосфатного пути начинается с дегидрирования глюкозо-6-фосфата при С-1, реакции, катализируемой глюкозо-6-фосфат-дегидрогеназой.
Фермент высокоспецифичен в отношении NADP + ; Км для NAD + примерно в тысячу раз выше, чем для NADP + . Продуктом реакции является 6-фосфоглюконо-δ-лактон, внутримолекулярный эфир, с эфирной связью между С-1-карбоксильной группой и гидроксилом при С-5. Следующий этап — гидролиз 6-фосфоглюконо — δ — лактона специфической лактоназой, дающий 6-фосфо-глюконат. Этот шестиуглеродный сахар подвергается затем окислительному декарбоксилированию 6-фосфоглюконат — дегидрогеназой с образованием рибулозо-5-фосфата. Акцептором электронов вновь служит NADP + . (смотри рисунок). Конечным этапом синтеза рибозо-5-фосфата является изомеризация рибулозо-5-фосфата фосфопентозо-изомеразой. Эта реакция подобна гликолитическим реакциям:
Все три кетозо-альдозные изомеризации идут через образование ендиольного промежуточного продукта.
Окислительная фаза пентозофосфатного пути иногда считается основной и неокислительная рассматривается как связка с гликолизом. Это связано с тем фактом, что эти фазы могут идти как независимо друг от друга, так вместе. В ходе неокислительной фазы пентозофосфатного пути происходит регенерация рибозо-5-фосфата в глюкозо-6-фосфат. Основную роль в этом процессе играют два фермента: транскетолаза и трансалъдолаза. Эти же ферменты создают обратимую связь между пентозофосфатным путем и гликолизом, катализируя следующие три реакции:
Транскетолаза переносит двухуглеродную группу, включающую 1-й и 2-й атомы углерода кетозы, на альдегидный углерод альдозного сахара. Происходит, следовательно, превращение кетосахара в альдозу, содержащую на два атома углерода меньше, и одновременное превращение альдосахара в кетозу, содержащую на два атома углерода больше. Коферментом реакции является тиаминидифосфат (в его состав входит тиамин — витамин группы В), для ее протекания необходимы ионы Mg 2+ . Переносимая двухуглеродная группа является, вероятно, гликоальдегидом, связанным с тиаминдифосфатом, т. е. «активным гликольальдегидом». Транскетолаза катализирует перенос двухуглеродной группы с ксилулозо-5-фосфата на рибозо-5-фосфат с образованием семиуглеродной кетозы седогептулозо-7-фосфата и альдозы глицеральдегид-3-фосфата. Эти два продукта далее вступают в следующую реакцию, называемую трансальдолазной. Трансальдолаза осуществляет перенос трехуглеродного фрагмента, «активного дигидроксиацетона» (атомы углерода 1 — 3), кетозы седогептулозо-7-фосфата на альдозу глицеральдегид-3-фосфат; в результате образуются кетоза фруктоза-6-фосфат и четырехуглеродная альдоза эритрозо-4-фосфат. Следующая реакция снова катализируется транскетолазой. В этой реакции ксилулозо-5-фосфат служит донором «активного гликоальдегида». Роль акцептора выполняет образовавшийся ранее эритрозо-4-фосфат. Продуктами этой реакции являются фруктоза-6-фосфат и глицеральдегид-3-фосфат.
Итак, избыток рибозо-5-фосфата, образованный в пентозофосфатном пути, может количественно превращаться в промежуточные продукты гликолиза.
Значение метаболического пути для различных тканей можно оценить по его активности. Пентозофосфатный путь активно протекает в печени, жировой ткани, коре надпочечников, щитовидной железе, эритроцитах, семенниках и в молочных железах в период лактации; он неактивен в нелактирующей молочной железе и малоактивен в скелетных мышцах. Все ткани, в которых активность данного пути высока, используют в реакциях восстановительного синтеза NADPH, например в реакциях синтеза жирных кислот, стероидов, аминокислот (с участием глутаматдегидрогеназы) или восстановленного глутатиона в эритроцитах. Вероятно, в условиях активного липогенеза или при наличии любой системы, утилизирующей NADPH, возрастает активная деградация глюкозы по пентозофосфатному пути в связи с увеличением отношения NADP + /NADPH. В условиях, которые возникают после приема пищи, может индуцироваться синтез глюкоза-6-фосфатдегидрогеназы и 6-фосфоглюконатдегидрогеназы.
Читайте также: Текстиль дата мебельные ткани остатки
Регуляция скорости функционирования пентозофосфатного пути
Первая реакция окислительной ветви пентозофосфатного пути, дегидрирование глюкозо-6-фосфата, по существу необратима. Действительно, при физиологических условиях эта реакция лимитирует скорость процесса и выполняет функцию «контрольного пункта». Наиболее важным регуляторным фактором является концентрация NADP + , акцептора электронов при окислении глюкозо-6-фосфата в 6-фосфоглюконо-лактон. Кроме того, NADPH конкурирует с NADP + за связывание с ферментом, и АТР конкурирует с глюкозо-6-фосфатом. Отношение концентрации NADP + к концентрации NADPH в цитозоле печени крыс, содержащихся на полноценном рационе, составляет примерно 0,014, что на несколько порядков ниже отношения [NAD + ] / [NADH], которое при этих же условиях равно 700. Выраженное действие концентрации NADP + на скорость превращений по окислительной ветви пентозофосфатного пути подтверждает, что генерирование NADPH тесно сопряжено с его использованием в восстановительных биосинтезах. Вопрос о регуляции неокислительной ветви пентозофосфатного пути до сих пор остается открытым.
Регуляция направления пентозофосфатного шунта
Судьба глюкозо-6-фосфата зависит от потребности в NADPH, рибозо-5-фосфате и АТФ.
В данном случае возможно четыре различные ситуации (схема регуляторных механизмов представлена на рисунке 5).

Рисунок 5: Схема регуляции направлений пентозофосфатного шунта
1. Потребность в рибозо-5-фосфате значительно превышает потребность в NADPH. Большая часть глюкозо-6-фос-фата превращается во фруктозо-6-фостфат и глицеральдегид-3-фосфат по гликолитическому пути. Затем две молекулы фрукто-зо-6-фосфата и одна молекула глицераль-дегид-3-фосфата превращаются под действием трансальдолазы и транскетолазы в три молекулы рибозо-5-фосфата путем обращения реакции, описанной ранее. Стехиометрия этого превращения следующая:
Пентозофосфатный путь окисления глюкозы
Для начала нужно понять, что это за путь такой и какая молекула в него вступает. Пентозофосфатный путь — это путь окисления глюкозы. Точнее, не совсем глюкозы, а глюкозо-6-фосфата. Мы уже знаем, что глюкоза, которая попадает в клетку, фосфорилируется гексокиназой или глюкокиназой. В зависимости от типа ткани. В печени работает глюкокиназа, а в остальных тканях — гексокиназа. Клетка получает глюкозо-6-фосфат, который можно отправить на синтез гликогена или окислить до пирувата с помощью гликолиза.
Но существует другой путь окисления глюкозо-6-фосфата — пентозофосфатный. Если в ходе гликолиза клетка стремится получить энергию из этой молекулы, то в пентозофосфатном у неё другие цели. И вот какие: получить кофермент НАДФH и пентозы. И тут вы задаётесь вопросом: «Неужели эти молекулы так важны, что для них существует целый отдельный путь?». Да, важны — сейчас посмотрим почему.
Функции пентозофосфатного пути
Мы первый раз встречаемся с коферментом НАДФH, до этого речь шла только о НАДH. Этот восстановленный кофермент используется для некоторых химических реакций, но его основная роль — отдать два электрона в дыхательную цепь. В дыхательной цепи из электронов получится 2,5 молекулы АТФ. Но я отвлёкся.
НАД + отличается от НАДФ + всего на одну фосфатную группу, но при этом у них огромное различие в функциях.
Для чего же клетке нужен НАДФH?
- Синтеза жирных кислот;
- Синтеза холестерола;
- Синтеза нейромедиаторов;
- Образования дезоксирибозы — компонента нуклеотидов ДНК;
- Реакций обезвреживания ксенобиотиков и активных форм кислорода.
Довольно большой список. Даже если ты ещё не встречался с этими процессами, то он всё равно впечатляет. Видно, что у НАДФH огромная роль в нашем организме. И вот в чём прикол: пентозофосфатный путь почти единственный способ получить НАДФH для клеточек. Есть ещё малик-фермент и дегидрирование изоцитрата, но о них в другой раз. В норме клетке хватает пентозофосфатного пути для синтеза НАДФH.
Но это только первый продукт пентозофосфатного пути, а ведь были ещё пентозы. А если точнее, то в ходе пентозофосфатного пути образуется рибоза-5-фосфат. Она используется для синтеза:
- Нуклеотидов ДНК и РНК;
- АТФ;
- Коферментов НАД, НАДФ, ФАД и А.
У всех этих молекул есть кое-что общее. В их формулах есть рибоза, либо дезоксирибоза. А дезоксирибоза это производное чего? Рибозы. Респект.
Пентозофосфатный путь — поставщик НАДФH и пентоз для клетки.
Кажется, что эти функции противоположны двум первым, но это не так. У пентозофосфатного пути есть пластичность. В конце статьи мы поговорим о ней, а пока обсудим классический вариант.
Реакции пентозофосфатного пути
Реакции протекают в цитоплазме клетки, там же где и гликолиз. Пентозофосфатный путь состоит из двух этапов: окислительного и неокислительного. Мне не совсем нравится слово «этап». Оно подразумевает, что это два друг за другом следующих процесса. Но к концу статьи вы поймёте, что пентозофосфатный путь может идти не только последовательно, но и отдельно по каждому этапу. Ну вот, я опять забежал вперёд.
Читайте также: Какими способами можно закрепить конец нитки в ткани ответ
Окислительный этап
Мы договорились, что в пентозофосфатный путь вступает глюкозо-6-фосфат. В ходе окислительного этапа из этой молекулы образуется рибулозо-5-фосфат. Глюкоза теряет один атом углерода — из гексозы образуется пентоза. Давайте прикинем, что нужно для этого. Легко отщепить атом углерода, когда он карбоксильный — ведь у нас есть реакции декарбоксилирования. В них отщепляется кислотная группа COOH. В глюкозо-6-фосфате, увы, такой группы нет. Есть только альдегидная, да и та участвует в образовании циклической формы (вспомнить про альдозы и кетозы можно тут). Поэтому окислительный этап будет направлен на раскрытие цикла и перевод альдегидной группы в карбоксильную.
Первая реакция. Катализатор — глюкозо-6-фосфатдегидрогеназа. Кофермент реакции НАДФ. В ходе этой реакции от молекулы глюкозо-6-фосфата отщепляется два водорода. При этом НАДФ присоединяет к себе гидрид ион. Гидрид ион состоит из протона водорода и двух электронов. Образуется глюконолактон-6-фосфат и НАДФH. Один протон отправляется в цитоплазму.
Для того, чтобы превратить глюконолактон-6-фосфат в пентозу нужно раскрыть цикл. Ведь наша молекула до сих пор в циклической форме. За счёт дегидрирования мы увеличили нестабильность связи, с помощью которой образуется цикл. Теперь эту связь легко порвать.
Вторая реакция. Фермент — глюконолактон-гидратаза. У неё есть название попроще — лактоназа. Лактоназа присоединяет молекулу воды по нестабильной связи, что приводит к её разрушению. В ходе этой реакции образуется 6-фосфоглюконат.
Эти две реакции нужны были для раскрытия шестичленного цикла. Но не только. Ещё мы превратили альдегидную группу глюкозо-6-фосфата, которая находится у первого углерода, в карбоксильную группу. Или превратили альдегид в кислоту. Ведь реакция декарбоксилирования пойдёт только при наличии карбоксильной группы. Да-да, я капитан очевидность.
Третья реакция. Катализирует реакцию 6-фосфоглюконатдегидрогеназа, а её кофермент — НАДФ. В этой реакции происходят два важных момента: глюконат теряет карбоксильную группу, и от пентозы отщепляются два водорода. Продукты реакции — рибулозо-5-фосфат и НАДФH.
На этом заканчивается окислительный этап. Клетка синтезировала рибулозо-5-фосфат из глюкозо-6-фосфата. Она превратила шестиуглеродную альдозу в пятиуглеродную кетозу. И не менее важно, что клетка получила две молекулы НАДФH.
Итог окислительного этапа: образование пятиуглеродной кетозы и двух НАДФH
И ещё одно — все эти реакции были необратимыми. Этим окислительный этап отличается от неокислительного. Там все реакции будут обратимыми.
Неокислительный этап
Четвёртая реакция. Далее из рибулозо-5-фосфата можно получить рибозо-5-фосфат. Для этого нужно провести реакцию изомеризации. Ведь рибоза — это альдоза, а не кетоза. Фермент — фосфопентозоизомераза. Или просто изомераза.
Образуется рибозо-5-фосфат. Клетка получила то, что хотела — рибозо-5-фосфат. Теперь его можно пустить на синтез ДНК, РНК, АТФ, НАД, НАДФ, ФАД или кофермента А. Это в идеале.
Что, если клетке не нужны эти молекулы? Допустим, что сейчас ей всего достаточно — она укомплектована по полной. Но при этом ей нужен НАДФ. Клетка хочет синтезировать жирные кислоты или ей нужно что-то обезвредить. А двух молекул НАДФH как-то маловато. Что делать?
Можно сделать вот как. Пустим больше молекул глюкозо-6-фосфата на пентозофосфатный путь — получим больше НАДФH. Но при этом в клетке накапливаются пентозы, которые ей не нужны. Невыгодно. Глупо тратить глюкозо-6-фосфат, получая только половину плюшек.
Поэтому клетка делает по-другому. Она превращает рибозо-5-фосфат обратно в глюкозу. Но как? Ведь ты сказал, что окислительный этап необратим. А я и не врал, для этого нужен неокислительный этап. Вообще суть неокислительного этапа — перебрасывание углеродов с одной молекулы на другую. Сорри за спойлеры.
Вернёмся к третьей реакции. В ходе неё был получен рибулозо-5-фосфат. Давайте предположим, что мы пустили на пентозофосфатный путь три молекулы глюкозо-6-фосфата, а не одну. Не думай, почувствуй — дальше всё станет понятно. Клетка получит три молекулы рибулозо-5-фосфата. Из них одну мы превратим в рибозо-5-фосфат. А две другие в ксилулозо-5-фосфат. Их тоже будет два.
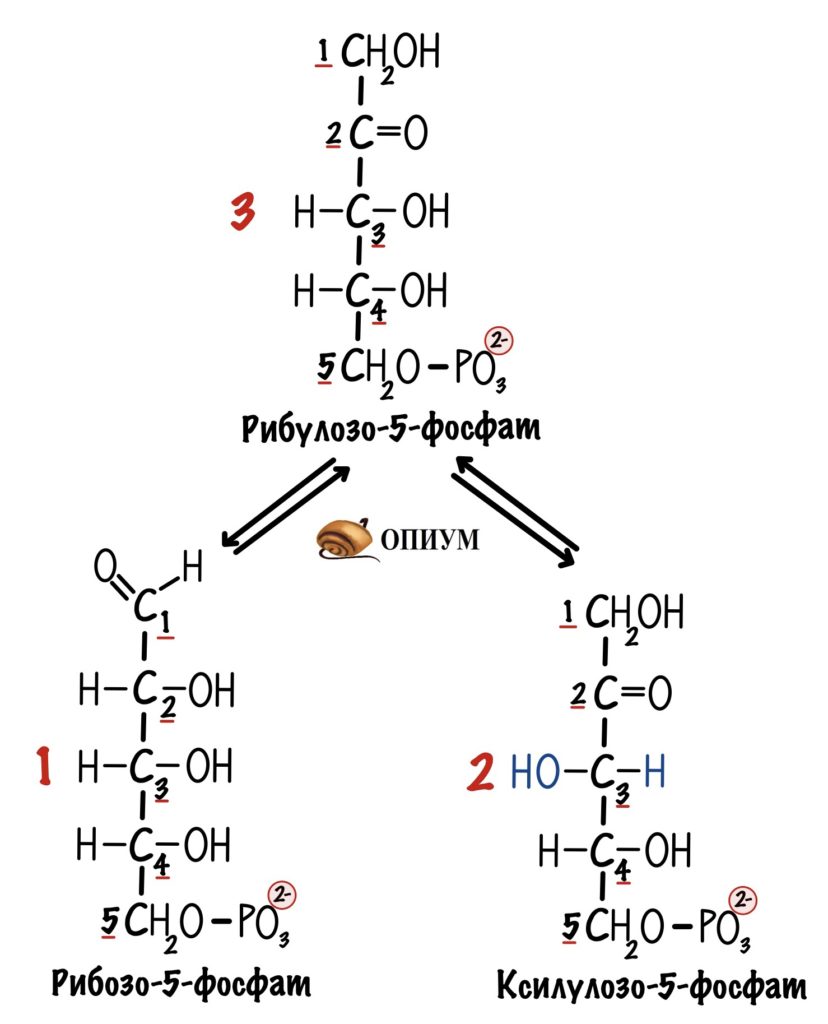
Фермент — фосфопентозоэпимераза. Или просто эпимераза. Она изменяет положение гидроксильной группы у третьего атома углерода. Теперь она слева, а не справа.
Можно начать синтезировать глюкозу из двух ксилулозо-5-фосфатов и одного рибозо-5-фосфата. Это невозможно сделать в одну реакцию, поэтому клетке понадобится целая серия. Сначала два атома углерода с ксилулозо-5-фосфата переносятся на рибозо-5-фосфат. Катализирует реакцию транскетолаза, она не может работать без своего кофермента — тиаминдифосфата.
Читайте также: Поперечнополосатая ткань количество ядер
В ходе этой реакции получаем глицеральдегид-3-фосфат и седогептулозо-7-фосфат (какое сложное название). Первый образовался из ксилулозо-5-фосфата. Для образования альдегидной группы нужна была реакция эпимеризации — превращения рибулозо-5-фосфата в ксилулозо-5-фосфат. А второй из рибозо-5-фосфата. Клетка получает триозу и гептозу.
Продолжаем перекидывать атомы углерода с одной молекулы на другую. Теперь у нас другой фермент — трансальдолаза. Она переносит три атома углерода с седогептулозо-7-фосфата на глицеральдегид-3-фосфат.
Образуется фруктозо-6-фосфат и эритрозо-4-фосфат. Вроде всё неплохо, фруктозо-6-фосфат можно превратить в глюкозу, но что делать с эритрозо-4-фосфатом? Вспоминаем, что у нас есть ещё одна молекула ксилулозо-5-фосфата. Снова используем транскетолазу. Она перекидывает два атома углерода с ксилулозо-5-фосфата на эритрозо-4-фосфат. Получаем ещё один фруктозо-6-фосфат и глицеральдегид-3-фосфат.
Получается, что из трёх молекул рибулозо-5-фосфата были синтезированы две молекулы фруктозо-6-фосфата и один глицеральдегид-3-фосфат. Из двух фруктозо-6-фосфатов образуется два глюкозо-6-фосфата.
А вот общая схема неокислительного этапа:
Глицеральдегид-3-фосфат — знакомая молекула, она встречалась нам в гликолизе. Это значит, что её можно отправить в глюконеогенез. Правда для того, чтобы синтезировать молекулу глюкозы нужно два глицеральдегид-3-фосфата. Ну давайте решим эту проблему тем, что удвоим количество глюкозо-6-фосфата, который был пущен на пентозофосфатный путь.
Мы взяли три молекулы, а теперь возьмём шесть. Из них будет синтезировано шесть молекул рибулозо-5-фосфата. Превратим их в две молекулы рибозо-5-фосфата и четыре молекулы ксилулозо-5-фосфата. Так мы получим четыре фруктозо-6-фосфата и два глицеральдегид-3-фосфата. Из фруктозо-6-фосфата образуется четыре молекулы глюкозо-6-фосфата, а две молекулы глицеральдегид-3-фосфата превратим в ещё один фруктозо-6-фосфат. Получается, что общее уравнение такого пентозофосфатного пути будет:
6 глюкозо-6-фосфат + 12НАДФ + + 6H2O = 5 глюкозо-6-фосфат + 12НАДФH + 12H + + 6CO2.
Поздравляю! Мы превратили наш пентозофосфатный путь в цикл. Пусть он и не будет работать слишком долго, но так мы уменьшим потери глюкозо-6-фосфата на образование НАДФH.
Пластичность пентозофосфатного пути
Ну а теперь посмотрим на пластичность пентозофосфатного пути в разных ситуациях.
Ситуация 1. Клетке нужны НАДФH и пентозы.
Допустим, что клетка готовится к делению. Ей нужно реплицировать ДНК и создать запас питательных веществ. На синтез ДНК нужны пентозы, а на запас — НАДФH.
Здесь всё просто — отправляем глюкозо-6-фосфат на окислительный этап. Далее превращаем рибулозо-5-фосфат в рибозо-5-фосфат. При этом получаем две молекулы НАДФH и одну пентозу. Использовать по потребности.
Ситуация 2. Клетке нужен НАДФH, но не нужны пентозы.
Такая ситуация будет в эритроцитах. В норме гемовое железо имеет заряд +2, но существуют метгемоглобинобразователи, которые переводят его в +3. При этом образуются активные формы кислорода. Им не место в эритроците — они могут повредить его мембрану. Повреждение мембраны приведёт к гемолизу. Вот мы и потеряли эритроцит. Но в эритроците есть герой — глутатион. Это трипептид, у него имеется -SH группа. Глутатион восстанавливает активные формы кислорода, отдавая электроны. А сам окисляется. Для того, чтобы вернуть глутатион в активное состояние нужен фермент глутатионпероксидаза. Она переносит электроны с НАДФH на глутатион, восстанавливая его. Вот такая сложная система.
Проводим окислительный этап, а далее зацикливаем процесс. Превращаем шесть молекул рибулозо-5-фосфата в пять молекул глюкозо-6-фосфата. Крутим, пока клетке не надоест этим заниматься. Эту ситуацию мы разбирали выше.
Ситуация 3. Клетке нужны пентозы, но не нужен НАДФH.
В клетке нет кофермента НАДФ + , но делиться всё-таки надо! Так что придётся выкручиваться из этой ситуации.
Забываем про окислительный этап. Помните, что в неокислительном этапе все реакции обратимы? Это значит, что мы можем повернуть их в другую сторону. В ходе гликолиза получаем четыре молекулы фруктозо-6-фосфата. Далее используем трансальдолазные и транскетолазные реакции для того, чтобы получить пять молекул рибозо-5-фосфата.
Правда для этого нам понадобится две молекулы глицеральдегид-3-фосфата, которые мы так же получим в ходе гликолиза. Но не трогая четыре фруктозо-6-фосфата, которые мы уже взяли до этого. Для этого пустим ещё одну молекулу глюкозо-6-фосфата в гликолиз.
Ситуация 4. Клетке нужны НАДФH и АТФ.
Такая ситуация тоже может быть в эритроцитах, когда им не нужно много НАДФH.
В этом случае будем отправлять глюкозо-6-фосфат на окислительный этап. Получаем пентозы и НАДФH. Пентозы с помощью транскетолазных и трансальдолазных реакций превращаем в фруктозо-6-фосфат и глицеральдегид-3-фосфат. Отправляем их в гликолиз. В эритроцитах идёт только анаэробный гликолиз, поэтому получаем лактат. В других клетках гликолиз будет аэробным — образуется пируват. Синтезированный пируват пойдёт в цикл трикарбоновых кислот.
Хочешь задать вопрос, похвалить или наговорить гадостей? Тогда залетай в телегу. Там ты сможешь предложить новый формат или разбор темы. А если серьёзно, то эти статьи пишутся для вас, поэтому мне важна обратная связь.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
