
При повреждении нервов дегенеративные процессы аксона происходят дистальнее места поражения. Это связано с тем, что аксон является отростком нейрона, и его питание и рост зависят от сомы. Регенеративные процессы периферической нервной системы характеризуются высокой интенсивностью и в большинстве случае обеспечивают полное восстановление структур. В ЦНС, напротив, интенсивность процессов восстановления не высока, и регенерация не является полной.
а) Валлерова (антероградная) дегенерация периферических нервов. Основные этапы процесса дегенерации и их описание представлены на рисунке ниже и в подписи к нему. После разрушения или перерезки нерва в течение первых 48 ч аксоны и миелиновая оболочка под действием протеаз, высвобождающихся из шванновских клеток под влиянием ионов Са 2+ , распадаются на фрагменты элипсоидной формы дистальнее места нарушения целостности нерва. Продукты распада удаляют моноциты, которые перемещаются из кровотока в эндоневрий и превращаются в макрофаги.
Помимо осуществления фагоцитоза, макрофаги стимулируют митотическую активность шванновских клеток. Макрофаги и шванновские клетки выполняют трофическую (питательную) и направляющую функции для регенерирующих аксонов.
В исходе дегенеративного процесса формируется «бугристый» цитоскелет нерва, в котором соединительная ткань и периневрий остаются сохранными, окружая делящиеся шванновские клетки.
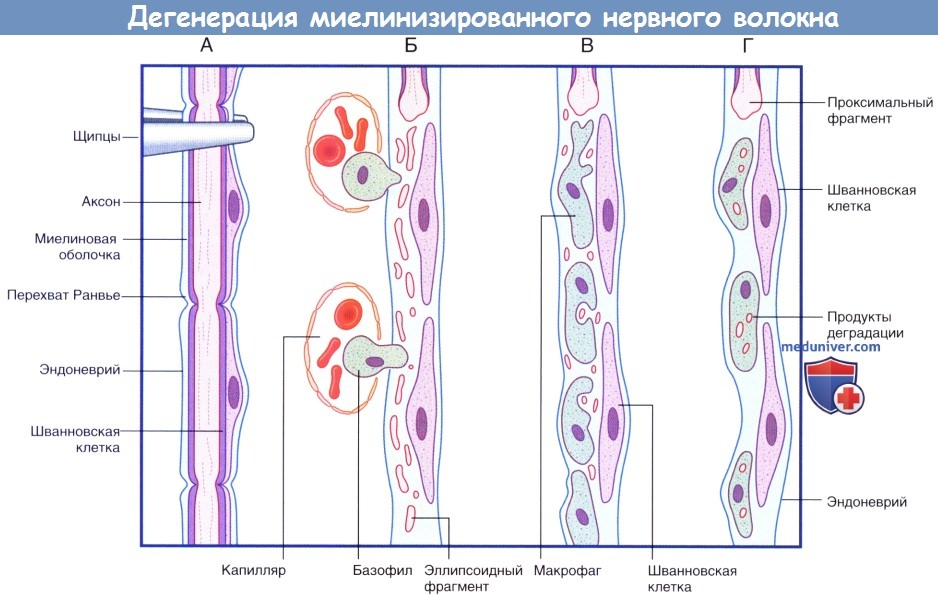 Последовательность процессов при дегенерации миелинизированного нервного волокна.
Последовательность процессов при дегенерации миелинизированного нервного волокна.
(А) Сохранное нервное волокно; продемонстрированы четыре сегмента. Выполняют пережатие нервного волокна в верхней части.
(Б) Миелиновая оболочка и аксон распадаются на мелкие частицы и фрагменты эллипсоидной формы. Моноциты проникают в эндоневрий из кровеносного русла.
(В) Мелкие продукты дегенерации поглощают моноциты.
(Г) Происходит практически полное удаление всех продуктов дегенерации. Шванновские клетки и эндоневрий сохранны. Последовательность процессов при регенерации миелинизированного нервного волокна. 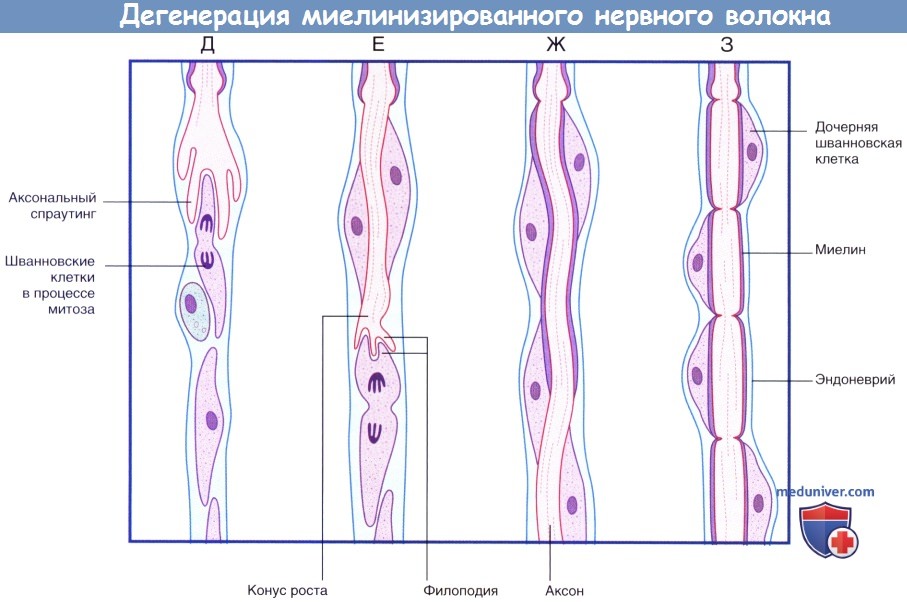 (продолжение) Последовательность процессов при дегенерации миелинизированного нервного волокна.
(продолжение) Последовательность процессов при дегенерации миелинизированного нервного волокна.
(Д) Аксональный спраутинг начинается с дистального участка аксона. Спраутинг оказывает митогенное дейсвтие на окружающие шванновские клетки.
(Е) Конус роста продвигается дистально вдоль поверхности шванновских клеток.
(Ж) Миелинизация начинается вдоль проксимального участка регенерирующего аксона.
(З) Общее строение вновь миелинизированного нервного волокна соответствует тем же принципам, однако миелинизированные сегменты характеризуются меньшей длиной.
б) Регенеративные процессы в периферической нервной системе. Основные этапы процесса регенерации представлены на рисунке ниже. После ровного среза нерва спраутинг (разрастание новых ветвей) на конце проксимального отрезка аксона начинается уже спустя несколько часов. Однако в клинической практике повреждения нерва часто происходят при раздавливании или разрыве. В этих случаях происходит отмирание участка нерва длиной 1 см и более, за счет чего спраутинг может продолжаться в течение недели. В случае удачной регенерации происходит тесное соприкосновение проксимального конца аксона со шванновской клеткой дистального конца пересеченного нейрона.
При нарушении формирования этой связи в месте первичного повреждения образуется псевдоневрома, представляющая собой извитые регенерирующие аксоны, погруженные в рубцовую ткань. Ампутационные псевдоневромы — источники сильных болей после ампутаций конечностей.
Регенерация нейронов при повреждении происходит двумя путями в течение нескольких часов после повреждения. На проксимальном конце пересеченного аксона появляются множественные отростки, на конце которых образуются утолщения — конусы роста. На дистальном конце шванновские клетки формируют отростки, направляющиеся навстречу конусам роста. На концах конусов роста формируются напоминающие антенны филоподии, где располагаются поверхностные рецепторы, временно связывающиеся с соответствующими поверхностными молекулами адгезии базальных мембран шванновских клеток. Актиновые филаменты филоподий прикрепляются к поверхностным рецепторам и относительно этих соединений осуществляют дальнейшее продвижение конусов роста.
Конусы роста стимулируют митотическую активность шванновских клеток. Шванновские клетки делятся и миелинизируют наиболее крупные аксоны.
На начальном этапе регенерация аксонов у человека происходит со скоростью около 1 мм/сут., однако с течением времени регенеративная способность нейронов и поддерживающая функция дистально денервированных шванновских клеток снижаются. Например, в том случае, если восстановление иннервации концевой пластинки двигательного нейрона на мышце не произошло в течение 12 мес, восстановление функции данной структуры маловероятно. Кроме того, прогноз для восстановления функции более благоприятен при размозжении тканей, чем при полном нарушении целостности нерва, поскольку в случае размозжения эндоневрий остается сохранным. Филоподии двигательных и чувствительных аксонов «распознают» базальные мембраны тех шванновских клеток, которые до повреждения окружали соответствующий тип аксонов.
При полном разрыве нервных стволов перед попыткой их восстановления, как правило, выжидают около трех недель, поскольку сразу после повреждения их соединительнотканные оболочки отечны, а в течение этого промежутка времени они становятся немного толще, что позволяет шовному материалу лучше закрепиться. Кроме того, обрезание нервов перед наложением швов приводит к развитию вторичной аксотомии проксимальной части пересеченного аксона. В ходе экспериментальных исследований на животных показано, что вторичная аксотомия стимулирует более интенсивную и длительную регенерацию.
Читайте также: Ткань для летних платьев штапель
Влияния тела нейрона на пересеченный участок нерва заключаются в следующем:
• В течение нескольких дней после нарушения целостности аксона тельца Ниссля в материнских клетках спинального ганглия и серого вещества спинного мозга перестают окрашиваться катионными красителями. Этот феномен получил название хроматолизиса («обесцвечивания»). По данным электронной микроскопии отмечают увеличение количества шероховатой эндоплазматической сети и ее распространение по всему объему перикариона. Кроме того, шероховатая эндоплазматическая сеть образует скопления в глубине плазматической мембраны.
• Вследствие осмотических изменений перикариона ядро ориентируется эксцентрически.
• Клетки нейроглии заполняют все синаптические щели, изолируя исходный двигательный нейрон от синаптических контактов в сером веществе.
• В исследованиях, проведенных на обезьянах, показано, что после перерезки чувствительных нейронов 30-40 % окончаний дорсальных нервных корешков подвергается валлеровской дегенерации. Регенерация этих нейронов не происходит, поскольку их окончания располагаются в сером веществе головного мозга. Однако некоторые рецепторы восстанавливаются за счет спраутинга сохранных прилежащих нейронов. Данное наблюдение позволяет объяснить явление неполного восстановления чувствительности у таких пациентов.
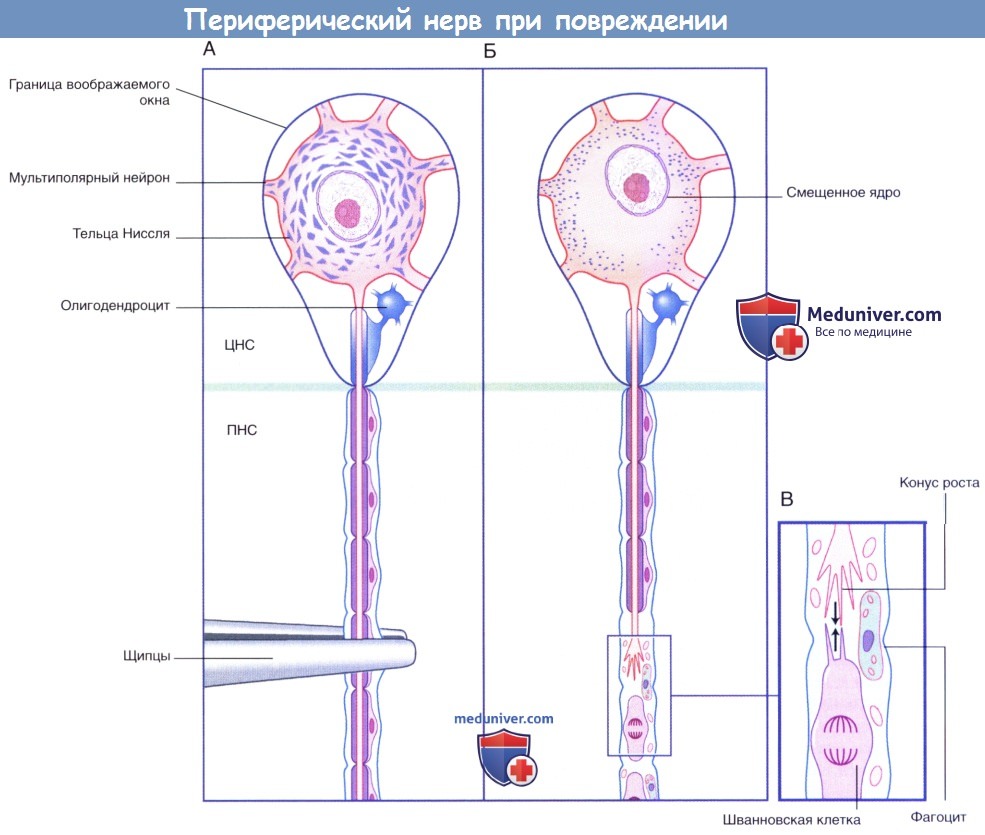
Схематическое изображение процессов, происходящих в периферическом нерве после повреждения.
(А) Двигательный нейрон ЦНС, видимый через воображаемое окно.
(Б) Хроматолизис проявляется фрагментированием и рассеиванием телец Ниссля, а также смещением ядра.
(В) В зоне повреждения в условиях удаления продуктов деградации происходит образование контакта филоподий конуса роста с проксимальными выростами шванновских клеток (указано стрелками).
ЦНС — центральная нервная система; ПНС — периферическая нервная система.
Редактор: Искандер Милевски. Дата публикации: 12.11.2018
Рост нервов в тканях
Введение. Проблема физиологической и репаративной регенерации нервной системы всегда была в центре внимания нейробиологов и неврологов. В конце ХХ века большое количество исследований было посвящено исследованию трансплантации нервной ткани. (Полежаев и др., 1993; Семченко и др., 2000). Однако, несмотря на определенные достижения, полноценного приживления нервной ткани и восстановления функциональных связей при разного видах алло- и ксенотрансплантациях достигнуть не удалось.
Открытие нейрональных стволовых клеток (НСК), их обнаружение во взрослой нервной системе позвоночных животных и человека и развитие клеточных технологий позволило по новому взглянуть на эту проблему (Семченко и др. 2012; К.Н. Ярыгин, В.Н. Ярыгин, 2012; Ярыгин и др., 2015). В данной работе приводится краткий обзор собственных и имеющихся в литературе данных по регенерации нервной ткани в норме и в условиях эксперимента.
Нейрональные стволовые клетки (НСК) относятся к группе тканеспецифичных или региональных стволовых клеток. Они обладают характеристиками самоподдерживающейся популяции клеток, которые при дифференцировке способны давать нейроны, астроциты и олигодендроциты в развивающемся и взрослом мозге. Впервые они были обнаружены в ЦНС человека в 1995 году при анализе посмертных срезов мозга, окрашенных иммунногистохимически на BrdU (бромдезоксиуридин) (Gage et al., 1995). В настоящее время найден целый ряд нейрональных маркеров, которые позволяют более или менее надежно идентифицировать НСК и их потомки. Среди них следует отметить: ядерный антиген нервных клеток – NeuN; маркер нейробластов – даблкортин (DCX); нейрон-специфическую энолазу – NSE; молекулы адгезии нервных клеток – PSA-NCAM; цитоскелетные белки – нестин, β-тубулин III; транскрипционные факторы – Sox-1, Sox-2, Dlx2, Pax 6 и ряд других. Часть из этих маркеров специфична для клеток нервной ткани, другие направлены на выявление свойств, характерных для разных популяций стволовых клеток. (Гомазков, 2014; Коржевский и др., 2010, 2015).
Обнаружение НСК как в развивающемся, так и во взрослом мозге поставило вопрос об их происхождении. В ранний период эмбрионального развития НСК происходят из клеток нейроэпителия, которые путем симметричного и асимметричного деления дают начало нейронам, глиальным клеткам и клеткам-предшественникам, которые включаются в процессы нейрогенеза на более поздних этапах развития, включая постнатальный период. Характер деления клеток – один из механизмов выбора НСК путей развития. В случае симметричного митоза образуются две одинаковые дочерние клетки, которые либо сохраняют пролиферативный потенциал – т.е. остаются стволовыми, либо могут уйти на путь нейрогенеза или глиогенеза – опять же обе. При втором варианте симметричного деления популяция стволовых клеток может потерять способность к самообновлению и истощится. При ассиметричном делении одна клетка остается пролиферативной (стволовой), другая выходит в дифференцировку. При этом пул НСК сохраняется.
Популяция нейрональных клеток – предшественников достаточно гетерогенна. На основную роль предшественника в позднем пренатальном и в постнатальном периодах претендует т.н. «радиальная глия – RG». Она выполняет двоякую роль – ее отростки, пронизывающие всю толщу развивающейся стенки нервной трубки, служат направляющими для миграции молодых нейробластов, а также обладает потенциями НСК, давая начало новым популяциям нейронов и глии. Особо необходимо отметить, что именно из потомков радиальной глии формируются основная масса интернейронов коры головного мозга млекопитающих животных и человека. Иммунологически в клетках радиальной глии помимо маркеров, традиционных для глиальных клеток (ГКФБ, виментин, нестин), выявляются специфические маркеры радиальной глии: фермент ароматаза–В (Aro-B), BLBP (brain lipid binding protein) и GLAST – глутаматный транспортер. Фермент ароматаза-В (Aro-B) связан с синтезом ароматизированных стероидов и синтезируется в клетках радиальной глии мозга молодых и взрослых позвоночных животных. Для них также характерна экспрессия полисиаловой молекулы адгезии нервных клеток (PSA-NCAM), транскриптационного фактора Sox-2 и фактора RG2. Важно подчеркнуть, что в клетках радиальной глии обнаруживаются маркеры и нейрональной линии дифференцировки (ТН- тирозингидроксилаза, ГАМК и NADP – диафораза). (Обухов, Пущина, 2011). На роль нейрональных предшественников также претендуют клетки эпендимы, астроциты, NG2 клетки и танициты (Гомазков, 2014; Pinto, Götz. 2007).
Читайте также: Неактивный илеит с гиперплазией лимфоидной ткани
Таким образом, во взрослом мозге позвоночных животных и человека сохраняются группы клеток-предшественников, которые обладают свойствами НСК и способны в течение длительного периода обновлять популяции нейронов и глии.
Нейрогенные ниши. Нейрональные стволовые клетки и их потомки находятся в тесном взаимодействии со многими элементами окружающей их структуры мозга, формируя вмести с ними своеобразную – нейрогенную нишу (stem niche). В ее состав входят НСК и их потомки, клетки эпендимы, астроциты, олигодендроциты, эндотелий капилляров мозга и компоненты межклеточного матрикса (Обухов и др., 2015). Клетки «ниши» способны экспрессировать целый ряд факторов, необходимых для сохранения популяции НСК и регуляции нейро- и глиогенеза. Среди факторов, влияющих на нейрогенез следует отметить группу транскрипционных факторов (Shh, Sox1, Sox2, Tbr1, Wnt, BMP, Notch1, Pax6 и др). Они действуют на разные стадии нейро- и глиогенеза, причем часто прямо противоположно. Например: транскрипционные факторы Notch1 и BMP подавляют нейрональную дифференцировку, направляя развитие клеток предшественников в глиальном направлении, а фактор Trb2 наоборот – стимулирует нейрогенез. Фактор Shh (sonic hedgehog) и транскрипционные факторы из семейства Sox регулируют процесс пролиферации клеток-предшественников. Эти факторы могут влиять и на другие этапы нейро-и глиогенеза, проходящие в пролиферативных зонах (миграцию нейробластов, образование определенных типов клеток, формирование отростков у нейронов, развитие синаптических связей). Разнообразные ростовые факторы (эпидермальный фактор роста – EGF, трансформирующий фактор роста – TGFa, основной фактор роста фибробластов – bFGF, инсулиноподобный ростовой фактор – IGF1, фактор роста эндотелия – VEGF; интерферон гамма – IFN-γ и др.) также влияют на пролиферацию и дифференцировку клеток-предшественников. Особое место среди сигнальных молекул занимают нейромедиаторы и нейромодуляторы. В настоящее время установлено, что нейроны вскоре после образования из клеток-предшественников и задолго до начала миграции и формирования межнейрональных связей начинают секретировать молекулы нейромедиаторов, которые оказывают существенное влияние на развитие клеток в течение эмбриогенеза, а также в ходе постэмбрионального нейрогенеза. В последнее время особое внимание уделяется роли газообразных посредников (NO, H2S, CO) в регуляции процессов нейрогенеза в пре- и постнатальном периодах развития ЦНС. Было показано, что они существенно влияют на процесс миграции нейробластов, рост аксонных и дендритных ветвлений, пролиферацию и апоптоз НСК и их потомков в нейрогенных нишах и за их пределами (Пущина и др., 2012; Яковлев, Ситдикова, 2014; Puschina et al., 2011, 2014).
Особо следует отметить, что сами НСК способны синтезировать и секретировать подобные вещества, действующие в данном случае по типу пара- или аутокринной регуляции.
Организация пролиферативных зон в мозге млекопитающих. Зоны взрослого нейрогенеза у млекопитающих, включая приматов, обнаружены в субвентрикулярной зоне (SVZ) латеральных мозговых желудочков конечного мозга и в субгранулярной зоне (SGZ) зубчатой фасции гиппокампа. Наличие подобных зон в других отделах ЦНС млекопитающих в настоящее время не доказано, а имеющиеся данные носят крайне противоречивый характер. (Ярыгин и др., 2014; Ярыгин, Ярыгин, 2012). Субвентрикулярная зона (SVZ) образована несколькими слоями клеток (от двух до пяти), в составе которых выделяют несколько типов клеток. Скорость увеличения числа новых клеток в зубчатой извилине гиппокампа (SGZ) взрослого мозга определяется как 9000 единиц в течение суток, что составляет примерно 6 % от ощего количества нейронов в зубчатой фасции гиппокампа крысы.или около 250 тысяч в месяц. (Cameron, McKay, 2001). Вновь образованные нервные клетки мигрируют на места своей локализации в данной структуре мозга, формируют систему отростков и синапсов и встраиваются в функциональные нейронные сети. Cледует отметить, что, хотя факт интеграции новых нейронов в существующие нейронные сети доказан, функциональные аспекты этого процесса во многом еще неясны.
Читайте также: Шью из лоскутков мебельной ткани
В этом плане весьма интересным является обнаружение подобных пролиферативных зон в разных отделах головного мозга у представителей других групп позвоночных животных (рыб, амфибий, птиц). Взрослый нейрогенез у этих животных идет более интенсивно и дольше, чем у млекопитающих. (Обухов и др., 2015; Puschina et al., 2014; Grandel, Brand, 2013).
Взрослый нейрогенез и перспективы репаративной регенерации нервной ткани. Исследование последствий ишемии мозга показали, что она сопровождается усилением нейрогенеза в пролиферативных зонах и миграцией молодых клеток в зону повреждения. (Гомазков, 2014; Solway et al., 1998). Эти данные вызвали целую серию работ, направленных на изучение возможности использования НСК и их потомков для трансплантации в поврежденный мозг, а также поиска модельных объектов для экспериментальных работ (Григорьян, Кругляков, 2008; Семченко и др., 2012). Одной из удачных моделей явились рыбы разных видов. В серии работ с помощью иммуногистохимического маркирования PCNA (пролиферативного ядерного антигена), ядерного маркера нейрональной дифференцировки (HuCD), транскриптационного фактора Pax6 и серии других маркеров, в разных отделах мозга рыб был идентифицирован ряд пролиферативных зон (ПВЗ), свидетельствующих о наличии постоянного постнатального нейрогенеза в ЦНС рыб. (Обухов и др., 2015; Zupanc, 2009; Zupanc, Sîrbulescu, 2013; Puschina, Obukhov, 2012; Pushchina et al., 2014 a, b). Однако, в настоящее время неизвестно как этот процесс связан с нейрогенезом во взрослом мозге, и какие элементы матричных зон мозга рыб участвуют в репаративном нейрогенезе.
Были поставлены эксперименты на молоди нескольких видов рыб, которым наносили механическую травму в разные структуры мозга (зрительный нерв, крыша среднего мозга, полушария конечного мозга). При механическом повреждении разных отделов мозга молоди рыб (сетчатки, среднего мозга и мозжечка) было выявлено усиление пролиферативной активности как в традиционных пролиферативных зонах нейрогенеза (перивентрикулярные области), так зафиксировано появление новых нейрогенных участков. Процесс репарации после нанесения механической травмы глаза начинается с апоптоза поврежденных элементов. Апоптотический ответ наблюдается уже через полчаса после нанесения повреждающего воздействия и продолжается до 21 дня после нанесения травмы. Эти данные подтверждены результатами маркирования TUNEL-позитивных фрагментов ДНК в зоне повреждения (зрительного нерва), а также данными электронно-микроскопического анализа. Ультраструктурные изменения ядра свидетельствуют о различных стадиях процесса апоптоза в поврежденных клетках. Апоптоз, как механизм элиминации поврежденных в результате травмы клеток мозга рыб существенно отличается от такового у млекопитающих. У последних, основным способом элиминации поврежденных клеток в зоне травмы является некроз. Апоптоз же затрагивает незначительный объем клеток в прилегающих к травме областях. Наличие некроза в зоне травмы млекопитающих является одной из причин развития последующего вторичного воспаления в зоне повреждения, что в свою очередь вызывает дальнейшее нарастание некротического ответа в области травмы, в результате которого формируются большие полости, лишённые клеток. Эти полости, как правило, ограничены зоной реактивных астроцитов, создающих как механический, так и биохимический барьеры, затрудняющие рост нервных волокон и миграцию клеток в зону повреждения. В отличие от некроза при апоптозе отсутствуют признаки воспалительной реакции, а сами клетки впоследствии уничтожаются с помощью макрофагов/микроглии. Прижизненный мониторинг клеток в зоне повреждения с помощью мультифотонной конфокальной микроскопии показал, что уже через час после повреждающего воздействия наблюдается физиологический ответ со стороны макрофагов и микроглии, которые мигрируют в область нанесения механической травмы и активно участвуют в элиминации поврежденных клеток с помощью фагоцитоза. Дифференцировка клеток в нейрональном направлении, обнаруженная при помощи маркирования клеток антителами против белка HuC/D, происходила в пролиферативных зонах теленцефалона, зрительного тектума, мозжечка и продолговатого мозга форели уже через 2 дня после травмы (Пущина и др., 2016; Puschina et al., 2014).
Таким образом, показано, что после механической травмы в мозге экспериментальных животных источником новых нейронов являются появляющиеся в пролифератиных областях мозга новые зоны индуцированного нейрогенеза: нейрогенные ниши и участки вторичного нейрогенеза. (Пущина и др., 2016). Полученные данные послужат основой для дальнейших исследований особенностей постнатального нейрогенеза в ЦНС животных и человека в норме и при патологии.
Работа выполнена при финансовой поддержке гранта Президента РФ (МД 4318.2015.04) и Программы фундаментальных исследований ДВО РАН «Дальний Восток» (проект № 15-I-6-116).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом