
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
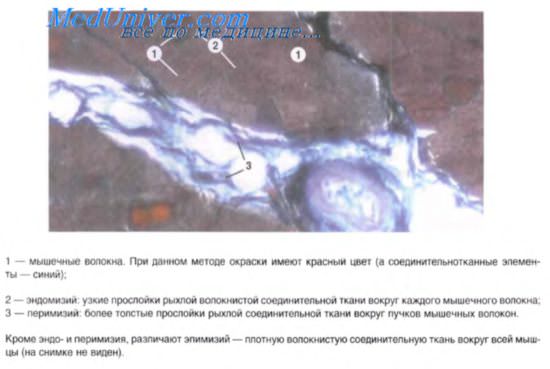
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Читайте также: Как сделать бабочку галстук своими руками из ткани без машинки
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
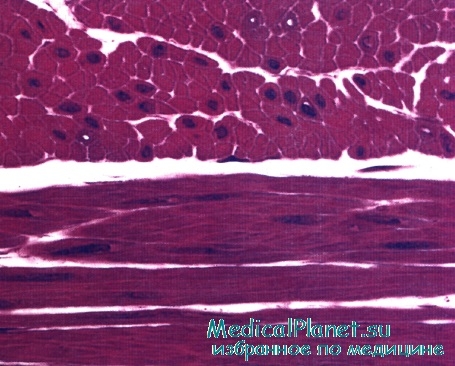
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
Сердечная мышечная ткань строение регенерация
Гистогенез сердечной мышечной ткани. Источники развития сердечной мышечной ткани находятся в прекардиальной мезодерме. В гистогенезе возникают парные складчатые утолщения висцерального листка спланхнотома — миоэпикардиальные пластинки, содержащие стволовые клетки сердечной мышечной ткани. Последние путем дивергентной дифференцировки дают начало следующим клеточным дифферонам: рабочим, ритмзадающим (пейсмекерным), проводящим и секреторным кардиомиоцитам.
Исходные клетки сердечной мышечной ткани — кардиомиобласты характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндоплазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролиферативным потенциалом. После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез. В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постнатальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов. Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофибриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов «конец в конец», «конец в бок» формируют клеточные комплексы — сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру.
Строение сердечной мышечной ткани.
Структурно-функциональные единицы волокон — кардиомиоциты — это клетки, имеющие вытянутую прямоугольную форму. Длина рабочих кардиомиоцитов составляет 50-120 мкм, а ширина — 15-20 мкм. Одно-два ядра располагаются в центре клетки. Периферическую часть цитоплазмы кардиомиоцитов занимают поперечноисчерченные миофибриллы, аналогичные таковым в симпластах скелетномышечного волокна. Однако каналы саркоплазматической сети и Т-системы менее отчетливо выражены. Кардиомиоциты отличаются большим количеством митохондрий, расположенных тесными рядами между миофибриллами. Снаружи миоциты покрыты сарколеммой, в составе которой выделяются плазмолемма и базальная мембрана. Характерной особенностью ткани является наличие вставочных дисков на границе между контактирующими кардиомиоцитами. Вставочные диски пересекают волокно в виде волнистой или ступенчатой линии и включают межклеточные контакты от простых, по типу десмо-сом и до щелевых (нексусов).
Читайте также: Ткань для вещь мешка армейского
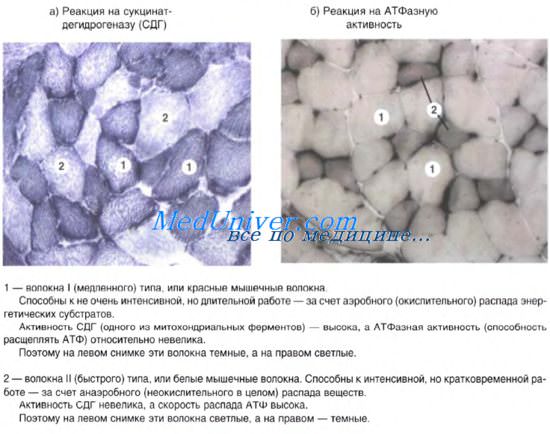
Часть кардиомиоцитов на ранних этапах кардиомиогенеза являются сократительно-секреторными. В дальнейшем в результате дивергентной дифференцировки возникают «темные» (сократительные) и «светлые» (проводящие) миоциты, в которых исчезают секреторные гранулы, тогда как в предсердных миоцитах они сохраняются. Так формируется дифферон эндокринных кардиомиоцитов. Эти клетки содержат центрально расположенное ядро с диспергированным хроматином,
1-2 ядрышками. В цитоплазме хорошо развиты гранулярная эндоплазматическая сеть, диктиосомы комплекса Гольджи, в тесной связи с элементами которого находятся многочисленные секреторные гранулы диаметром около 2 мкм, содержащие электронноплотный материал. В дальнейшем секреторные гранулы обнаруживаются под сарколеммой и выделяются в межклеточное пространство путем экзоцитоза. Выделенный пептидный гормон кардиодилатин циркулирует в крови в виде кардионатрина, который вызывает сокращение гладких миоцитов артериол, увеличение почечного кровотока, ускоряет клубочковую фильтрацию и выделение натрия из организма.
Кардиомиоциты проводящей системы гетероморфны. В них слабо развит мио-фибриллярный аппарат, расположение миофиламентов в составе миофибрилл рыхлое, Z-линии имеют неправильную конфигурацию, эндоплазматическая сеть слабо развита, находится на периферии миоцитов, число митохондрий незначительное. По мере расположения этих кардиомиоцитов в проксимо-дистальном направлении соответственно движению импульсов от синусно-предсердного узла, через предсердно-желудочковый узел, пучок Гиса, его ножки и клетки Пуркиня к рабочим миоцитам проводящие кардиомиоциты по своей ультраструктуре приближаются к рабочим кардиомиоцитам.
Регенерация сердечной мышечной ткани.
В гистогенезе сердечной мышечной ткани специализированный камбий не возникает. Поэтому регенерация ткани протекает на основе внутриклеточных гиперпластических процессов. Вместе с тем для кардиомиоцитов млекопитающих, приматов и человека характерен процесс полиплоидизации. Например, у обезьян ядра до 50% терминально дифференцированных кардиомиоцитов становятся тетра- и октоплоидными. Полиплоидные кардиомиоциты возникают за счет ацитокинетического митоза, что приводит к многоядерности.
В условиях патологии сердечно-сосудистой системы человека (ревматизм, врожденные пороки сердца, инфаркт миокарда и другие) важная роль в компенсации повреждений кардиомиоцитов принадлежит внутриклеточной регенерации, полиплоидизации как ядер, так и кардиомиоцитов.
Структурно-функциональная характеристика сердечной мышечной ткани. Источники развития и регенерации.
Собственно сердечная, мышечная ткань по своим свойствам занимает промежуточное положение между гладкими мышцами внутренних органов и поперечнополосатыми (скелетными). Она сокращается быстрее гладких, но медленнее поперечнополосатых мышц, работает ритмично и мало утомляется.
В связи с этим в ее строении имеется ряд своеобразных черт .Состоит эта ткань из отдельных мышечных клеток (миоцитов), почти прямоугольной формы, расположенных столбиком друг за другом. В целом получается структура, напоминающая поперечнополосатое волокно, разделенное на отрезки поперечными перегородками — вставочные диски,. Рядом лежащие волокна соединены анастомозами, что позволяет им сокращаться одновременно.
Структурно-функциональной единицей является клетка —кардиомиоцит. По строению и функциям кардиомиоциты подразделяются на две основные группы:
· типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
Читайте также: Как убрать пятна с белых кроссовок из ткани
· атипичные кардиомиоциты составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности. Синусныекардиомиоциты передают управляющие сигналыпереходнымкардиомиоцитам, а последние-проводящим
Есть также .секреторные кардиомиоциты вырабатываютнатрийуретический фактор
Гистогенез Источники развития поперечно-полосатой мышечной ткани-симметричные участки висцерального листка спланхнотома в шейной части зародыша—миоэпикардиальные пластинки.Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 5 видов кардиомиоцитов- рабочие, синусные переходные, проводящие, а также секреторные.
Возможности регенерации сердечной мышечной ткани: Стволовых клеток в сердечной мышце нет, поэтому погибающие кардиомиоциты не восстанавливаются.
Гладкая мышечная ткань. Структурная организация разновидностей гладких мышечных тканей. Структурные основы сокращения гладких мышечных клеток.. Иннервация.
Гладкая мышечная ткань образует достаточно большое количество важных внутренних органов в животном теле. Так, ей образованы: кишечник; половые органы; кровеносные сосуды всех типов; железы; органы выделительной системы; дыхательные пути; части зрительного анализатора; органы пищеварительной системы. –
Различают три группы гладких мышечных тканей- мезенхимные, эпидермальные и нейральные.
Структурная основа -Гладкий миоцит- веретеновидная клетка. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается.
. Соседние миоцитов через отверстия в базальной мембране образуют друг с другом щелевидные сообщения (нексус), которые обеспечивают функциональные взаимодействия клеток.
Сократительных аппарат гладких миоцитов (миофибрилл) состоит из тонких миофиламентов, образованных актином, и толстым, сформированным миозином. Миоциты ограничены базальной мембраной, а также коллагеновыми (ретикулярными) эластичными волокнами
Эфферентная (моторная) иннервация гладких миоцитов осуществляется постганглионарными волокнами автономной нервной системы
Сокращение ГМТ медленное — тоническое, зато ГМТ малоутомляема.
ГМТ в эмбриональном периоде развивается из мезенхимы
Нервные ткани. Источники развития. Синапсы. Классификация синапсов.Строение, механизмы передачи импульсов
Нервная ткань— это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и передачи его.
Она является основой строения органов нервной системы, обеспечивающих регуляцию всех тканей и органов, их интеграцию в организме и связь с окружающей средой.
Нервные клетки(нейроны, нейроциты) — основные структурные компоненты нервной ткани, выполняющие специфическую функцию.
Нейроглия(neuroglia) обеспечивает существование и функционирование нервных клеток, осуществляя опорную, трофическую, разграничительную, секреторную и защитную функции.
Развитие. Нервная ткань развивается из дорсальной эктодермы. У 18-дневного эмбриона человека эктодерма формируетнервную пластинку, латеральные края которой образуютнервные валики, а между валиками формируется нервныйжелобок. Передний конец нервной пластинки образует головной мозг. Латеральные края образуютнервную трубку.Полость нервной трубки сохраняется у взрослых в виде системы желудочков головного мозга и центрального канала спинного мозга.
Синапсы– это структуры, предназначенные для передачи импульса с одного нейрона на другой или на мышечные и железистые структуры.
В зависимости от способа передачи импульсасинапсы могут быть химическими или электрическими (электротоническими).
Химические синапсы передают импульс на другую клетку с помощью специальных биологически активных веществ — нейромедиаторов, находящихся в синаптических пузырьках.
Область синаптического контакта между двумя нейронами состоит из пресинаптической мембраны, синаптической щели и постсинаптической мембраны.
Электрические, или электротонические, синапсы в нервной системе млекопитающих встречаются относительно редко. В области таких синапсов цитоплазмы соседних нейронов связаны щелевидными соединениями (контактами), обеспечивающими прохождение ионов из одной клетки в другую,
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом