

Главная / Полезная информация / Статьи / Кардиология
Несмотря на то, что сердечная мышца имеет авторитмичность, частота сердечных сокращений модулируется эндокринной и нервной системами.
Существуют два основных типа сердечно-мышечных клеток : миокардиальные сократительные клетки и клетки миокарда.
По сравнению с гигантскими цилиндрами скелетных мышц, клеток сердечной мышцы или кардиомиоцитов значительно короче с гораздо меньшими диаметрами. Сердечная мышца также демонстрирует страты, чередующийся узор тёмных полос А и светлых полос I, приписываемых точному расположению миофиламентов и фибрилл, которые организованы в саркомерах по длине клетки. Эти сократительные элементы практически идентичны скелетной мышце. T (поперечные) канальцы проникают из поверхностной плазматической мембраны, сарколемма, внутрь клетки, позволяя электрическому импульсу достигать внутренней поверхности. Т-канальцы встречаются только на дисках Z, тогда как в скелетных мышцах они встречаются на стыке полос А и I. Таким образом, в сердечной мышце имеется около половины Т-канальцев, как у скелетных мышц. К тому же, саркоплазматический ретикулум хранит несколько ионов кальция, поэтому большая часть ионов кальция должна поступать извне клеток. Результатом является более медленное начало сокращения. Митохондрии многочисленны, обеспечивая энергию для сокращений сердца. Как правило, кардиомиоциты имеют одно центральное ядро, но в некоторых клетках могут находиться два или более ядра.
Клетки сердечной мышцы свободно расходятся. Соединение между двумя соседними ячейками обозначено критической структурой, называемой интеркалированным диском, что помогает поддерживать синхронизированное сжатие мышцы. Сарколемы из соседних клеток связываются вместе на интеркалированных дисках. Они состоят из десмосом, специализированных связываний протеогликанов, плотных соединений и большого количества щелевых соединений, которые позволяют проходить ионы между клетками и помогают синхронизировать сжатие. Межклеточная соединительная ткань также помогает связывать клетки вместе. Важность сильного связывания этих клеток связана с силами, оказываемыми сокращением.
Сердечная мышца претерпевает аэробные формы дыхания , в первую очередь, метаболизируя липиды и углеводы. Миоглобин, липиды и гликоген все хранятся в цитоплазме. Клетки сердечной мышцы подвергаются дерматическим сокращениям с длинными рефрактерными периодами, за которыми следуют короткие периоды релаксации. Релаксация необходима, поэтому сердце может заполнить кровью для следующего цикла. Огнеупорный период очень длинный, чтобы предотвратить возможность тетании, состояние, при котором мышца остаётся непроизвольно сжатой. В сердце, тетания несовместима с жизнью, так как это предотвратило бы сердце от перекачивания крови.
Повреждённые клетки сердечной мышцы имеют чрезвычайно ограниченные возможности для восстановления себя или замены мёртвых клеток через митоз. Недавние данные свидетельствуют о том, что, по крайней мере, некоторые стволовые клетки остаются в сердце, которые продолжают делить и потенциально заменять эти мёртвые клетки.
Сердечную мышечную ткань образуют три популяции кардиомиоцитов
Гистогенез сердечной мышечной ткани. Источники развития сердечной мышечной ткани находятся в прекардиальной мезодерме. В гистогенезе возникают парные складчатые утолщения висцерального листка спланхнотома — миоэпикардиальные пластинки, содержащие стволовые клетки сердечной мышечной ткани. Последние путем дивергентной дифференцировки дают начало следующим клеточным дифферонам: рабочим, ритмзадающим (пейсмекерным), проводящим и секреторным кардиомиоцитам.
Исходные клетки сердечной мышечной ткани — кардиомиобласты характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндоплазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролиферативным потенциалом. После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез. В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постнатальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов. Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофибриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов «конец в конец», «конец в бок» формируют клеточные комплексы — сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру.
Строение сердечной мышечной ткани.
Структурно-функциональные единицы волокон — кардиомиоциты — это клетки, имеющие вытянутую прямоугольную форму. Длина рабочих кардиомиоцитов составляет 50-120 мкм, а ширина — 15-20 мкм. Одно-два ядра располагаются в центре клетки. Периферическую часть цитоплазмы кардиомиоцитов занимают поперечноисчерченные миофибриллы, аналогичные таковым в симпластах скелетномышечного волокна. Однако каналы саркоплазматической сети и Т-системы менее отчетливо выражены. Кардиомиоциты отличаются большим количеством митохондрий, расположенных тесными рядами между миофибриллами. Снаружи миоциты покрыты сарколеммой, в составе которой выделяются плазмолемма и базальная мембрана. Характерной особенностью ткани является наличие вставочных дисков на границе между контактирующими кардиомиоцитами. Вставочные диски пересекают волокно в виде волнистой или ступенчатой линии и включают межклеточные контакты от простых, по типу десмо-сом и до щелевых (нексусов).
Часть кардиомиоцитов на ранних этапах кардиомиогенеза являются сократительно-секреторными. В дальнейшем в результате дивергентной дифференцировки возникают «темные» (сократительные) и «светлые» (проводящие) миоциты, в которых исчезают секреторные гранулы, тогда как в предсердных миоцитах они сохраняются. Так формируется дифферон эндокринных кардиомиоцитов. Эти клетки содержат центрально расположенное ядро с диспергированным хроматином,
Читайте также: Рисунок по ткани город
1-2 ядрышками. В цитоплазме хорошо развиты гранулярная эндоплазматическая сеть, диктиосомы комплекса Гольджи, в тесной связи с элементами которого находятся многочисленные секреторные гранулы диаметром около 2 мкм, содержащие электронноплотный материал. В дальнейшем секреторные гранулы обнаруживаются под сарколеммой и выделяются в межклеточное пространство путем экзоцитоза. Выделенный пептидный гормон кардиодилатин циркулирует в крови в виде кардионатрина, который вызывает сокращение гладких миоцитов артериол, увеличение почечного кровотока, ускоряет клубочковую фильтрацию и выделение натрия из организма.
Кардиомиоциты проводящей системы гетероморфны. В них слабо развит мио-фибриллярный аппарат, расположение миофиламентов в составе миофибрилл рыхлое, Z-линии имеют неправильную конфигурацию, эндоплазматическая сеть слабо развита, находится на периферии миоцитов, число митохондрий незначительное. По мере расположения этих кардиомиоцитов в проксимо-дистальном направлении соответственно движению импульсов от синусно-предсердного узла, через предсердно-желудочковый узел, пучок Гиса, его ножки и клетки Пуркиня к рабочим миоцитам проводящие кардиомиоциты по своей ультраструктуре приближаются к рабочим кардиомиоцитам.
Регенерация сердечной мышечной ткани.
В гистогенезе сердечной мышечной ткани специализированный камбий не возникает. Поэтому регенерация ткани протекает на основе внутриклеточных гиперпластических процессов. Вместе с тем для кардиомиоцитов млекопитающих, приматов и человека характерен процесс полиплоидизации. Например, у обезьян ядра до 50% терминально дифференцированных кардиомиоцитов становятся тетра- и октоплоидными. Полиплоидные кардиомиоциты возникают за счет ацитокинетического митоза, что приводит к многоядерности.
В условиях патологии сердечно-сосудистой системы человека (ревматизм, врожденные пороки сердца, инфаркт миокарда и другие) важная роль в компенсации повреждений кардиомиоцитов принадлежит внутриклеточной регенерации, полиплоидизации как ядер, так и кардиомиоцитов.
9.2.3. Сердечная мышечная ткань
Гистогенез и виды клеток. Источники развития сердечной поперечнополосатой мышечной ткани (textus muscularis striatus cardiacus) — симметричные участки висцерального листка спланхнотома в шейной части зародыша — миоэпикардиальные пластинки. Из них дифференцируются также клетки мезо-телия эпикарда. Исходные клетки сердечной мышечной ткани —кардиомио-бласты — характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндо-плазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролифератив-ным потенциалом.
После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез (рис. 9.9). В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постна-тальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов.
Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофи-бриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов «конец в конец», «конец в бок» формируют сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру. Часть кардиомиоцитов на ранних этапах кардиомиогенеза являются сократительно-секреторными. В дальнейшем в результате дивергентной дифференцировки возникают «темные» (сократительные) и «светлые» (проводящие) миоциты, в которых исчезают секреторные гранулы, тогда как в предсердных мио-цитах они сохраняются. Так формируется дифферон эндокринных кардиомиоцитов. Эти клетки содержат центрально расположенное ядро с диспергированным хроматином, одним-двумя ядрышками. В цитоплазме хорошо развиты гранулярная эндоплазматическая сеть, диктиосомы комплекса Гольджи, в тесной связи с элементами которого находятся многочисленные секреторные гранулы диаметром около 2 мкм, содержащие электронно-плотный материал. В дальнейшем секреторные гранулы обнаруживаются под сарколеммой и выделяются в межклеточное пространство путем экзоцитоза.
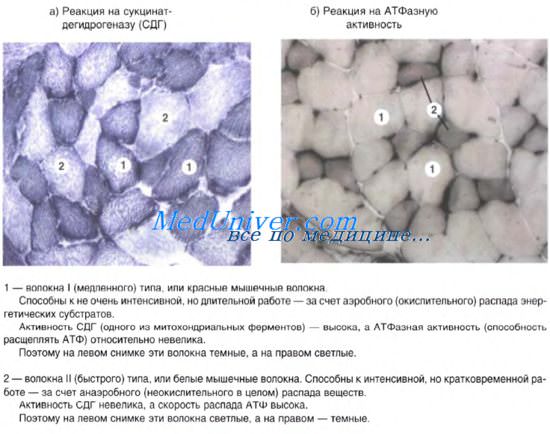
В целом в ходе гистогенеза возникает пять видов кардиомиоцитов — рабочие (сократительные), синусные (пейсмекерные), переходные, проводящие, а также секреторные. Рабочие (сократительные) кардиомиоциты образуют свои цепочки (рис. 9.10). Именно они, укорачиваясь, обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны

Рис. 9.9. Гистогенез сердечной мышечной ткани (схема по П. П. Румянцеву): а — кардиомиоциты в стенке сердечной трубки;б — кардиомиоциты в позднем эмбриогенезе; в — кардиомиоциты в постанатальном периоде. 1 — кардиомиоцит; 2 — митотически делящийся кардиомиоцит; 3 — миофиламенты и миофибриллы
передавать управляющие сигналы друг другу. Синусные (пейсмекерные) кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Клетки воспринимают управляющие сигналы от нервных волокон, в ответ на что изменяют ритм сократительной деятельности. Синусные (пейсмекерные) кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние — проводящим и рабочим кардиомиоцитам. Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами, и располагаются под эндо-
Читайте также: Что такое гистосовместимые ткани

Рис. 9.10. Строение сердечной мышечной ткани (микрофотография). Окраска — железный гематоксилин:
1 — ядро кардиомиоцита; 2 — цепочка кардиомиоцитов; 3 — вставочные диски
кардом. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее — другим проводящим кар-диомиоцитам. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим. Секреторные кардиомиоциты выполняют особую функцию. Они вырабатывают пептидный гормон кардиодилатин, который циркулирует в крови в виде кардионатрина, вызывает сокращение гладких миоцитов артериол, увеличение почечного кровотока, ускоряет клубочковую фильтрацию и выделение натрия. Все кардиомиоциты покрыты базальной мембраной.
Строение сократительных (рабочих) кардиомиоцитов. Клетки имеют удлиненную (100-150 мкм) форму, близкую к цилиндрической. Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна (толщиной до 20 мкм). В области контактов клеток образуются так называемые вставочные диски (рис. 9.10). Кардиомиоциты могут ветвиться и образуют пространственную сеть. Их поверхности покрыты базальной мембраной, в которую снаружи вплетаются ретикулярные и коллагеновые волокна. Ядро кардиомиоцита (иногда их два) овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения, за исключением агранулярной эндоплазматической сети и митохондрий.
Специальные органеллы, которые обеспечивают сокращение, называются миофибриллами. Они слабо обособлены друг от друга, могут расщепляться. Их строение аналогично строению миофибрилл миосимпласта скелетного мышечного волокна. Каждая митохондрия располагается на протяжении всего саркомера. От поверхности плазмолеммы в глубь кардиомиоцита направлены Т-трубочки, находящиеся на уровне Z-линии. Их мембраны сближены,
контактируют с мембранами гладкой эндоплазматической (саркоплазмати-ческой) сети. Петли последней вытянуты вдоль поверхности миофибрилл и имеют латеральные расширения (L-системы), формирующие вместе с Т-трубочками триады или диады (рис. 9.11, а). В цитоплазме имеются включения гликогена и липидов, особенно много включений миоглобина. Механизм сокращения кардиомиоцитов такой же, как у миосимпласта.
Организация кардиомиоцитов в ткань. Кардиомиоциты соединяются друг с другом по типу «конец в конец». Здесь образуются вставочные диски: эти участки выглядят как тонкие пластинки при среднем увеличении светового микроскопа. Фактически же концы кардиомиоцитов имеют неровную поверхность, поэтому выступы одной клетки входят во впадины другой. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмо-сомами (рис. 9.11, б).

Рис. 9.11. Строение кардиомиоцита: а — схема (по Ю. И. Афанасьеву и В. Л. Горячкиной); б — электронная микрофотография вставочного диска. Увеличение 20 000. 1 — миофибриллы; 2 — митохондрии; 3 — саркотубулярная сеть; 4 — Т-трубочки; 5 — базальная мембрана; 6 — лизосома; 7 — вставочный диск; 8 — десмосома; 9 — зона прикрепления миофибрилл; 10 — щелевые контакты; 11 — гликоген
К каждой десмосоме со стороны цитоплазмы подходит миофибрил-ла, закрепляющаяся концом в десмо-плакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами (щелевыми соединениями). Это создает между ними метаболические связи и обеспечивает синхронность сокращений.
Регенерация. В гистогенезе сердечной мышечной ткани камбий не возникает. Поэтому регенерация ткани протекает на основе внутриклеточных гиперпластических процессов. Вместе с тем для кардиомиоцитов млекопитающих, приматов и человека характерен процесс полиплоиди-
зации. Например, у обезьян ядра до 50 % терминально дифференцированных кардиомиоцитов становятся тетра- и октоплоидными. Полиплоидные кардиомиоциты возникают за счет ацитокинетического митоза, что приводит к многоядерности. В условиях патологии сердечно-сосудистой системы человека (ревматизм, врожденные пороки сердца, инфаркт миокарда и др.) важную роль в компенсации повреждений кардиомиоцитов играют внутриклеточная регенерация, полиплоидизация ядер, возникновение многоядерных кардиомиоцитов.
9.3. ГЛАДКИЕ МЫШЕЧНЫЕ ТКАНИ
Различают три группы гладких (неисчерченных) мышечных тканей (textus muscularis nonstriatus) и клеток: мезенхимные, нейральные и миоэпи-телиальные клетки.
9.3.1. Мышечная ткань мезенхимного происхождения
Гистогенез. Эта ткань делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоци-тов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их в цитоплазме становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние. Дифференцируясь, они синтезируют компоненты межклеточного матрикса, коллаген базальной мембраны, а также эластин. У дефинитивных клеток (миоцитов) синтетическая способность снижена, но не исчезает полностью.
Читайте также: Из чего делается ткань в расте
Строение и функционирование клеток. Гладкий миоцит — веретеновидная клетка длиной 20-500 мкм, шириной 5-8 мкм. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается (рис. 9.12-9.14).
Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерии обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм. Наибольшей длины гладкие мио-
циты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактильные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность.
Секреторные миоциты по своей ультраструктуре напоминают фиб-робласты, однако содержат в своей цитоплазме пучки тонких миофи-ламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцирован-ным. Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Эти участки хорошо видны на электронных микрофотографиях как плотные тельца. Мономеры миозина располагаются рядом с филаментами актина. Плазмолемма образует впя-чивания — кавеолы, в которых концентрируются ионы кальция. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их термина-лей, изменяет проницаемость плаз-молеммы. Происходит высвобождение ионов кальция, что влечет за собой как полимеризацию миозина, так и взаимодействие миозина с актином.
Происходит втягивание актино-вых миофиламентов между миози-
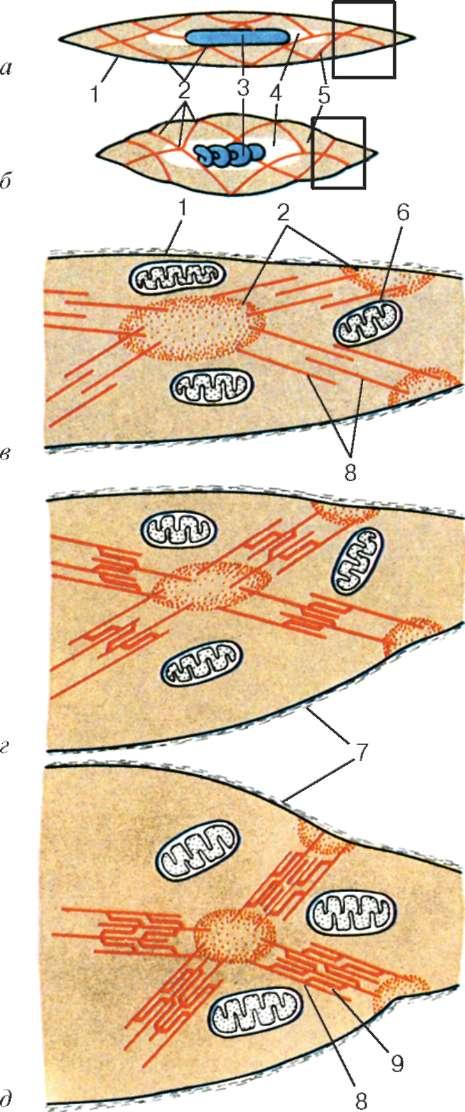
Рис. 9.12. Строение гладкого миоцита (схема):
а, в — при расслаблении; б, д — при наибольшем сокращении; г — при неполном сокращении; в-д — увеличенные изображения участков, обведенных рамками на фрагментах а и б. 1 — плазмолемма; 2 — плотные тельца; 3 — ядро; 4 — эндоплазма; 5 — сократительные комплексы; 6 — митохондрии; 7 — базальная мембрана; 8 — актиновые (тонкие) мио-филаменты; 9 — миозиновые (толстые) миофиламенты
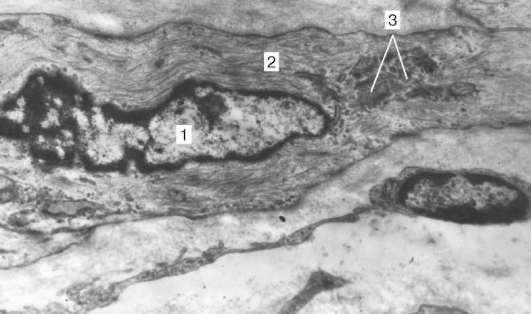
Рис. 9.13. Ультраструктура дифференцирующегося гладкого миоцита в стенке бронха:
1 — ядро; 2 — цитоплазма с миофиламентами; 3 — комплекс Гольджи, увеличение 35 000 (препарат А. Л. Зашихина)
новыми, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается (см. рис. 9.12). Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция перемещаются из цитоплазмы в кавеолы и в канальцы эндоплазматической сети, миозин деполимеризуется и «миофибриллы» распадаются. Сокращение прекращается. Таким образом, актиномиозиновые комплексы существуют в гладких миоцитах только в период сокращения при наличии в цитоплазме свободных ионов кальция.
Миоциты окружены базальной мембраной. На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс. Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и аморфный компонент межклеточного матрикса. Взаимодействие миоцитов осуществляется при помощи цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом, участков мембранных контактов поверхностей миоцитов.
Регенерация. Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов. В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазматического отношения, увеличение количества свободных рибосом и полисом, актива-

Рис. 9.14. Строение гладкой мышечной ткани (объемная схема) (по Р. В. Крстичу, с изменениями):
1 — веретеновидные гладкие миоциты; 2 — цитоплазма миоцита; 3 — ядра миоци-тов; 4 — плазмолемма; 5 — базальная мембрана; 6 — поверхностные пиноцитозные пузырьки; 7 — межклеточные соединения; 8 — нервное окончание; 9 — коллагеновые фибриллы; 10 — микрофиламенты
ция ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема). При действии ряда повреждающих факторов отмечается феноти-пическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании интимальной гиперплазии при развитии атеросклероза.

Рис. 9.15. Ультраструктура миопигментоцита (препарат Н. Н. Сарбаевой): 1 — ядро; 2 — миофиламенты, увеличение 6000
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
