
В самом общем понимании термин «синтез белка» означает несколько процессов, необходимых для преобразования гена в функциональный белок. Каждый процесс состоит из нескольких этапов, а регулирование может происходить на одном или нескольких этапах в рамках каждого процесса.
За последние 30 лет были достигнуты определенные успехи в понимании регуляции синтеза белка на молекулярном уровне, и теперь благодаря сведениям о полной последовательности генома у все большего числа различных организмов, в том числе грызунов и человека, новая информация, проясняющая и расширяющая наше понимание регуляции синтеза белка, накапливается быстрее, чем когда бы то ни было в прежние времена.
Обнаружение того, что геном человека содержит около 35 000 генов — примерно в 2 раза больше, чем было выявлено у беспозвоночных, — заставило научное сообщество пересмотреть влияние генома на разнообразие и сложность строения животного мира. В настоящее время принято считать, что движущей силой клеточного разнообразия является более глобальная регуляция синтеза белка.
Несомненно, у недоношенных и доношенных новорожденных движущей силой роста и развития является интенсификация синтеза белка. Основной прогресс в понимании того, каким образом нутритивные вещества (в частности, аминокислоты и глюкоза) регулируют синтез белка у недоношенных детей в возрасте до 1 мес, был достигнут с развитием модели новорожденных поросят.

Инфузия меченых аминокислот-маркеров в сочетании с разработанными в молекулярной биологии методами предоставила научному и медицинскому сообществу возможность более глубокого осмысления факторов, контролирующих накопление белков организмом. Кроме того, эти технологические достижения позволили исследовать влияние получаемого с питанием белка на отдельные ткани и органы в естественных условиях.
В эукариотических клетках способность экспрессировать биологически активные, или функциональные, белки попадает под основное регулирование по нескольким пунктам: транскрипция ДНК, обработка РНК, стабильность мРНК, трансляция мРНК и посттрансляционная модификация белка и его сворачивание.
Если бы содержание белков в клетке определялось на уровне транскрипции, взаимозависимость между уровнями белка и мРНК была бы линейной. На самом же деле связь между уровнями мРНК и содержанием белка в одной клетке является слабой, подчеркивая тот факт, что в регуляции содержания белка в клетке доминируют посттранскрипционные процессы. По этой причине двум последним пунктам регулирования (трансляция мРНК и посттрансляционные модификации белков и их сворачивание) в следующих статьях на нашем сайте уделено особое внимание.
![]()
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Синтез белков происходит в тканях
Белки, образуемые из полипептидов, — основные структурные компоненты тела человека, они катализируют большинство процессов метаболизма. Полипептидом называют цепь, состоящую из аминокислот, последовательность которых зависит от последовательности оснований в иРНК в соответствии с «генетическим кодом».
Каждая аминокислота представлена в иРНК одной или несколькими группа ми, называемыми триплетами кодонов, а их интерпретация в качестве полипептида носит имя трансляции. Трансляция иРНК происходит в рибосомах цитоплазмы в направлении от 5′- к З’-концу. Образуемые в результате полипептиды затем превращаются в белки.
Функциональные свойства белков обусловлены в основном их активными центрами, формирующимися при образовании третичной и четвертичной структур.
Генетический код для синтеза белка
Для трансляции необходимы молекулы тРНК, которые кодируют аминокислоты, соответствующие последовательности антикодона. Большинство аминокислот кодированы несколькими кодонами, исключение составляют триптофан и метионин (кодированы одним кодоном). Три из 64 возможных триплетных комбинаций А, Ц, Г и У в иРНК кодируют СТОП-сигналы: УГА, УАГ и УАА.
АУГ кодирует метионин и служит СТАРТ-сигналом, а также одновременно определяет амино-(N-)терминальный конец полипептида и формирует одну из трёх возможных рамок считывания. Генетический код митохондриальной ДНК немного отличается.
Трансляция при синтезе белка
Инициация. Малая субъединица рибосомы, содержащая несколько факторов инициации и кодирующая метионин метионил-тРНК, связывается с 5′-кэпом иРНК, а затем начинает скольжение вдоль молекулы иРНК и, достигнув первой АУГ-последовательности, сцепляется с ней. При этом происходит высвобождение факторов инициации, а большая субъединица рибосомы прикрепляется к малой, после чего запускается процесс трансляции.
Большая субъединица рибосомы имеет в своём составе два специализированных участка, известных как А-сайт (аминоацил-тРНК-связывающий участок) и Р-сайт (пептидил-тРНК-связывающий участок). В конце инициации к Р-сайту прикреплена заряженная мет-тРНК, антикодон которой сцеплен с АУГ-кодоном, в то время как А-сайт свободен.
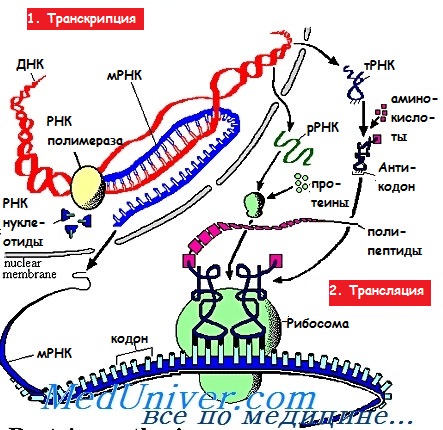
Элонгация. В соответствии с расположенным рядом кодоном иРНК к А-сайту при помощи растворимого фактора элонгации 1 (EF1) прикрепляется аминоацил-тРНК. Под воздействием пептидилтрансферазы возникает пептидная связь между аминогруппой (-NH2) аминокислоты на А-сайте и карбоксильной группой (-СООН) на Р-сайте, при этом первая тРНК отделяется.
Фермент транслоказа обеспечивает отделение свободной тРНК, движение рибосомы вдоль молекулы иРНК, а также перенос растущей пептидной цепочки от А-сайта к Р-сайту. Для осуществления данного процесса необходим фактор EF2.
Митохондриальные иРНК транслируются при помощи митохондриоспецифических тРНК.
Терминация. Элонгация продолжается до тех пор, пока на пути рибосомы не встретится один из терминирующих (СТОП) кодонов, которые распознают благодаря мультивалентным факторам освобождения (терминации, RF). При этом специфичность пептидилтрансферазы изменяется таким образом, что к | белку прикрепляется молекула воды. Затем рибосома отделяется и распадается на составляющие её субъединицы, освобождая готовый полипептид.
Для синтеза полипептида, длина которого составляет примерно 400 аминокислот, в среднем необходимо до 20 с.
Как только одна рибосома освобождает кэп иРНК, к нему присоединяется следующая, формируя полирибосому или полисому. Время жизни иРНК составляет несколько часов.
Строение белка
Аминокислотная последовательность полипептида обусловливает его первичную структуру.
Вторичная структура белка представляет собой трёхмерное расположение частей полипептида: а-спираль, коллагеновая про-а-спираль или бета-складочная конформация.
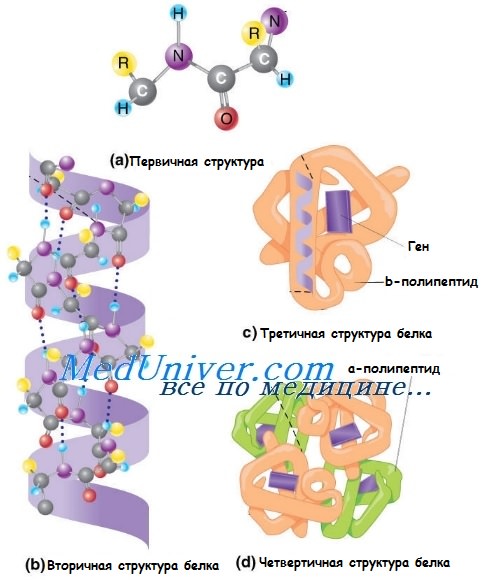
Третичная структура белка образуется при самоукладке всех вторичных структур полипептида.
Четвертичная структура белка — окончательная конформация мультимерного белка, такого, как, например, гемоглобин, который состоит из двух а-глобулиновых и двух бета-глобулиновых мономеров, а также молекулы гема и атома двухвалентного железа. Коллагеновые волокна состоят из трёхспиральных нитей, которые образованы тремя про-а-спиралями.
Чаще всего структуру белка поддерживают дисульфидные мосты, которые возникают между серосодержащими остатками цистеина расположенных рядом цепей, в то время как ферментативные свойства зависят прежде всего от распределения заряженных групп.
Посттрансляционные изменения белка
К посттрансляционным изменениям относят удаление N-терминального метионина и дробление. Объединение происходит между одинаковыми и разными полипептидами, а также между простетическими группами, к которым относят гем.
Полипептиды, предназначенные для внеклеточной (экстрацеллюлярной) секреции, вначале подвергаются гликозилированию в эндоплазматическом ретикулуме и аппарате Гольджи. Для их отбора имеет значение сигнальный пептид, расположенный около N-конца, который связывается с сигналраспознающей частицей, состоящей из цитоплазматической 7SL-PHK и шести специфических белков.
При помощи сигнального пептида происходит связывание с мембранным рецептором эндоплазматического ретикулума. Сразу после синтеза полипептид переносится через мембрану; как только появляется его С-конец (карбоксильная часть), сигнальный пептид отделяется. Затем полипептиды в составе везикул, которые отпочковываются от эндоплазматического ретикулума, попадают к аппарату Гольджи.
Читайте также: Для предотвращения омертвления тканей
Гликозилированию обычно подвержен N-конец пептида. При этом происходит присоединение олигосахарида к NH2-rpynne боковой цепи аспарагина (например, при производстве антител и лизоцима). Иногда олигосахарид присоединяется к ОН-группе боковой цепи серина, треонина или гидроксилизина (например, секретируемые антигены групп крови системы АВО).
Среди других модификаций различают гидроксилирование лизина и пролина, играющее важную роль в образовании про-а-спиралей коллагена, сульфатацию тирозина, служащую сигналом к компартментализации (пространственному разделению), а также липидизацию остатков цистеина и глицина, необходимую для их прикрепления к фосфолипидной мембране.
Ацетилирование лизина в гистоне Н4 влияет на его связывание с ДНК. Протеинкиназы фосфорилируют остатки серина и тирозина и могут регулировать ферментативную способность энзимов, как в случае с каскадной сигнальной трансдукцией протоонкогенов.
Медицинское значение понимания синтеза белка
В основе механизма возникновения болезни клеточных включений лежит недостаточность гликозилирования лизоцима. Рицин, входящий в состав бобов клещевины обыкновенной (Ricinus communis), блокирует EF2, а дифтерийный токсин — транслоказу.
Мишенью для большого количества антибиотиков служат процессы трансляции, характерные для прокариот. Например, эритромицин разрушает транслоказу, хлорамфеникол взаимодействует с пептидилтрансферазой, тетрациклин препятствует связыванию аминоацил-тРНК, пуромицин по своей структуре сходен с аминоацил-тРНК, а стрептомицин связывается с малой субъединицей бактериальных рибосом. При этом митохондрии человека имеют эволюционное родство с бактериями, поэтому некоторые антибиотики оказывают влияние и на митохондриальные функции.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Алгоритм метаболизма


автор: А. Ю. Барановский, д. м. н., профессор, заведующий кафедрой гастроэнтерологии и диетологии Северо-Западного государственного медицинского университета им. И. И. Мечникова, врач высшей категории
Решение организационных вопросов питания у лиц старших возрастов, разработка и назначение индивидуализированных рационов рационального, профилактического и лечебного питания в существенной степени зависит от правильной оценки врачом нутриционного статуса пожилого человека, особенностей состояния обменных процессов. Именно поэтому профессионально грамотный клиницист, участвующий в решении проблем лечебно-профилактического питания у лиц пожилого и старческого возраста, должен быть достаточно хорошо ориентирован в области основ клинической биохимии и физиологии питания стареющего организма.
Белковый обмен
Белки — сложные азотсодержащие биополимеры, мономерами которых служат аминокислоты (органические соединения, содержащие карбоксильные и аминные группы). Их биологическая роль многообразна. Белки выполняют в организме пластические, каталитические, гормональные, транспортные и другие функции, а также обеспечивают специфичность. Значение белкового компонента питания заключается прежде всего в том, что он служит источником аминокислот.
Аминокислоты делятся на эссенциальные и неэссенциальные в зависимости от того, возможно ли их образование в организме из предшественников. К незаменимым аминокислотам относятся гистидин, лейцин, изолейцин, лизин, метионин, фенилаланин, триптофан и валин, а также цистеин и тирозин, синтезируемые соответственно из метионина и фенилаланина. Девять заменимых аминокислот (аланин, аргинин, аспарагиновая и глутамовая кислоты, глутамин, глицин, пролин и серин) могут отсутствовать в рационе, так как способны образовываться из других веществ. В организме также существуют аминокислоты, которые продуцируются путем модификации боковых цепей вышеперечисленных (например, компонент коллагена — гидроксипролин — и сократительных белков мышц — 3-метилгистидин).
Большинство аминокислот имеют изомеры (D- и L-формы), из которых только L-формы входят в состав белков человеческого организма. D-формы могут участвовать в метаболизме, превращаясь в L-формы, однако утилизируются гораздо менее эффективно.
Взаимоотношение аминокислот
По химическому строению аминокислоты делятся на двухосновные, двухкислотные и нейтральные с алифатическими и ароматическими боковыми цепями, что имеет большое значение для их транспорта, поскольку каждый класс аминокислот обладает специфическими переносчиками. Аминокислоты с аналогичным строением обычно вступают в сложные, часто конкурентные взаимоотношения.
Так, ароматические аминокислоты (фенилаланин, тирозин и триптофан) близкородственны между собой. Хотя фенилаланин является незаменимой, а тирозин — синтезируемой из него заменимой аминокислотой, наличие тирозина в рационе как будто бы «сберегает» фенилаланин. Если фенилаланина недостаточно или его метаболизм нарушен (например, при дефиците витамина С) — тирозин становится незаменимой аминокислотой. Подобные взаимоотношения характерны и для серосодержащих аминокислот: незаменимой — метионина — и образующегося из него цистеина.
Триптофан в ходе превращений, для которых необходим витамин В 6 (пиридоксин), включается в структуру НАД и НАДФ, то есть дублирует роль ниацина. Приблизительно половина обычной потребности в ниацине удовлетворяется за счет триптофана: 1 мг ниацина пищи эквивалентен 60 мг триптофана. Поэтому состояние пеллагры может развиваться не только при недостатке витамина РР в рационе, но и при нехватке триптофана или нарушении его обмена, в том числе вследствие дефицита пиридоксина.
Аминокислоты также делятся на глюкогенные и кетогенные, в зависимости от того, могут ли они при определенных условиях становиться предшественниками глюкозы или кетоновых тел (см. табл. 1).
Таблица 1. Классификация аминокислот
| Виды | Эссенциальные аминокислоты | Неэссенциальные аминокислоты |
| Алифатические | Валин (Г), лейцин (К), изолейцин (Г, К) | Глицин (Г), аланин (Г) |
| Двухосновные | Лизин (К), гистидин (Г, К)* | Аргинин (Г)* |
| Ароматические | Фенилаланин (Г, К), триптофан (Г, К) | Тирозин (Г, К)** |
| Оксиаминокислоты | Треонин (Г, К) | Серин (Г) |
| Серосодержащие | Метионин (Г, К) | Цистеин (Г)** |
| Дикарбоновые и их амиды | Глутамовая кислота (Г), глутамин (Г), аспарагиновая кислота (Г), аспарагин (Г) | |
| Иминокислоты | Пролин (Г) |
Обозначения: Г — глюкогенные, К — кетогенные аминокислоты; * — гистидин незаменим у детей до года; ** — условно-незаменимые аминокислоты (могут синтезироваться из фенилаланина и метионина).
Необходимые азотсодержащие соединения
Поступление азотсодержащих веществ с пищей происходит в основном за счет белка и в менее значимых количествах — свободных аминокислот и других соединений. В животной пище основное количество азота содержится в виде белка. В продуктах растительного происхождения большая часть азота представлена небелковыми соединениями, также в них содержится множество аминокислот, которые не встречаются в организме человека и зачастую не могут метаболизироваться им.
Синтез пуриновых оснований
Человек не нуждается в поступлении с пищей нуклеиновых кислот. Пуриновые и пиримидиновые основания синтезируются в печени из аминокислот, а избыток этих оснований, поступивших с пищей, выводится в виде мочевой кислоты.
В синтезе пиримидиновых колец принимает участие витамин B 12 , для образования пуриновых структур необходима фолиевая кислота. Именно поэтому дефицит этих нутриентов отражается прежде всего на органе с высоким уровнем пролиферации, где идет наиболее интенсивный синтез нуклеиновых кислот, — на кроветворной ткани.
Прием белка
Обычный (но не оптимальный) ежедневный прием белка у среднестатистического человека составляет приблизительно 100 г. К ним присоединяется примерно 70 г белка, секретируемого в полость желудочно-кишечного тракта. Из этого количества абсорбируется около 160 г. Самим организмом в сутки синтезируется в среднем 240–250 г белка. Такая разница между поступлением и эндогенным преобразованием свидетельствует об активности процессов обратного восстановления исходного сложного химического соединения из «осколков», образовавшихся при его метаболизме (ресинтеза белков из аминокислот, а аминокислот из аммиака и «углеродных скелетов» аминокислот).
Читайте также: Габардин 100 полиэстер что за ткань
Азотное равновесие
Для здорового человека характерно состояние азотного равновесия, когда потери белка (с мочой, калом, эпидермисом и т. п.) соответствуют его количеству, поступившему с пищей. При преобладании катаболических процессов возникает отрицательный азотный баланс, который характерен для низкого потребления азотсодержащих веществ (низкобелковых рационов, голодания, нарушения абсорбции белка) и многих патологических процессов, вызывающих интенсификацию распада (опухолей, ожоговой болезни и т. п.). При доминировании синтетических процессов количество вводимого азота преобладает над его выведением, и возникает положительный азотный баланс, характерный для детей, беременных женщин и реконвалесцентов после тяжелых заболеваний.
После прохождения энтерального барьера белки поступают в кровь в виде свободных аминокислот. Следует отметить, что клетки слизистой оболочки желудочно-кишечного тракта могут метаболизировать некоторые аминокислоты (в том числе глутамовую кислоту и аспарагиновую кислоту в аланин). Способность энтероцитов видоизменять эти аминокислоты, возможно, позволяет избежать токсического эффекта при их избыточном введении.
Аминокислоты, как поступившие в кровь при переваривании белка, так и синтезированные в клетках, в крови образуют постоянно обновляющийся свободный пул аминокислот, который составляет около 100 г.
Путь белка
75 % аминокислот, находящихся в системной циркуляции, представлены аминокислотами с ветвящимися цепями (лейцином, изолейцином и валином). Из мышечной ткани в кровоток выделяются аланин, который является основным предшественником синтеза глюкозы, и глутамин. Многие свободные аминокислоты подвергаются трансформации в печени. Часть свободного пула инкорпорируется в белки организма и при их катаболизме вновь поступает в кровоток. Другие непосредственно подвергаются катаболическим реакциям. Некоторые свободные аминокислоты используются для синтеза новых азотсодержащих соединений (пурина, креатинина, адреналина) и в дальнейшем деградируют, не возвращаясь в свободный пул, в специфичные продукты распада.
Роль печени
Постоянство содержания различных аминокислот в крови обеспечивает печень. Она утилизирует примерно ⅓ всех аминокислот, поступающих в организм, что позволяет предотвратить скачки в их концентрации в зависимости от питания.
Первостепенная роль печени в азотном и других видах обмена обеспечивается ее анатомическим расположением — продукты переваривания попадают по воротной вене непосредственно в этот орган. Кроме того, печень непосредственно связана с экскреторной системой — билиарным трактом, что позволяет выводить некоторые соединения в составе желчи. Гепатоциты — единственные клетки, обладающие полным набором ферментов, участвующих в аминокислотном обмене. Здесь выполняются все основные процессы азотного метаболизма: распад аминокислот для выработки энергии и обеспечения глюконеогенеза, образование заменимых аминокислот и нуклеиновых кислот, обезвреживание аммиака и других конечных продуктов. Печень является основным местом деградации большинства незаменимых аминокислот (за исключением аминокислот с ветвящимися цепями).
Инсулиновый ответ
Синтез азотсодержащих соединений (белка и нуклеиновых кислот) в печени весьма чувствителен к поступлению их предшественников из пищи. После каждого приема пищи наступает период повышенного внутрипеченочного синтеза белков, в том числе альбумина. Аналогичное усиление синтетических процессов происходит и в мышцах. Эти реакции связаны прежде всего с действием инсулина, который секретируется в ответ на введение аминокислот и/или глюкозы.
Некоторые аминокислоты (аргинин и аминокислоты с ветвящимися цепями) усиливают продукцию инсулина в большей степени, чем остальные. Другие (аспарагин, глицин, серин, цистеин) стимулируют секрецию глюкагона, который усиливает утилизацию аминокислот печенью и воздействует на ферменты глюконеогенеза и аминокислотного катаболизма. Благодаря этим механизмам происходит снижение уровня аминокислот в крови после поступления их с пищей. Действие инсулина наиболее выражено для аминокислот, содержащихся в кровотоке в свободном виде (аминокислот с ветвящимися цепями), и малозначимо для тех, которые транспортируются в связанном виде (триптофана). Обратное инсулину влияние на белковый метаболизм оказывают глюкокортикостероиды.
Аминокислоты на «экспорт»
Печень обладает повышенной скоростью синтеза и распада белков по сравнению с другими тканями организма (кроме поджелудочной железы). Это позволяет ей синтезировать «на экспорт», а также быстро обеспечивать лабильный резерв аминокислот в период недостаточного питания за счет распада собственных белков.
Особенность внутрипеченочного белкового синтеза заключается в том, что он усиливается под действием гормонов, которые в других тканях производят катаболический эффект. Так, при голодании белки мышц, для обеспечения организма энергией, подвергаются распаду, а в печени одновременно усиливается синтез белков, являющихся ферментами глюконеогенеза и мочевинообразования.
Избыток белка и голодание
Прием пищи, содержащей избыток белка, приводит к интенсификации синтеза в печени и в мышцах, образованию избыточных количеств альбумина и деградации излишка аминокислот до предшественников глюкозы и липидов. Глюкоза и триглицериды утилизируются как горючее или депонируются, а альбумин становится временным хранилищем аминокислот и средством их транспортировки в периферические ткани.
При голодании уровень альбумина прогрессивно снижается, а при последующей нормализации поступления белка медленно восстанавливается. Поэтому хотя альбумин и является показателем белковой недостаточности, он низкочувствителен и не реагирует оперативно на изменения в питании.
7 из 10 эссенциальных аминокислот деградируют в печени — либо образуя мочевину, либо впоследствии используясь в глюконеогенезе. Мочевина преимущественно выделяется с мочой, но часть ее поступает в просвет кишечника, где подвергается уреазному воздействию микрофлоры. Аминокислоты с ветвящимися цепями катаболизируются в основном в почках, мышцах и головном мозге.
Роль мышц
Мышцы синтезируют ежедневно 75 г белка. У среднего человека они содержат 40 % от всего белка организма. Хотя белковый метаболизм происходит здесь несколько медленнее, чем в других тканях, мышечный белок представляет собой самый большой эндогенный аминокислотный резерв, который при голодании может использоваться для глюконеогенеза.
Мышцы являются основной мишенью воздействия инсулина: здесь под его влиянием усиливается поступление аминокислот, увеличивается синтез мышечного белка и снижается распад.
В процессе превращений в мышцах образуются аланин и глутамин, их условно можно считать транспортными формами азота. Аланин непосредственно из мышц попадает в печень, а глутамин вначале поступает в кишечник, где частично превращается в аланин. Поскольку в печени из аланина происходит синтез глюкозы, частично обеспечивающий мышцу энергией, получающийся круго- оборот получил название глюкозо- аланинового цикла.
К азотсодержащим веществам мышц также относятся высокоэнергетичный креатин-фосфат и продукт его деградации креатинин. Экскреция креатинина обычно рассматривается как мера мышечной массы. Однако это соединение может поступать в организм с высокобелковой пищей и влиять на результаты исследования содержания его в моче. Продукт распада миофибриллярных белков — 3-метилгистидин — экскретируется с мочой в течение короткого времени и является достаточно точным показателем скорости распада в мышцах — при мышечном истощении скорость его выхода пропорционально снижается.
Механизм голодания
В отсутствие пищи синтез альбумина и мышечного белка замедляется, но продолжается деградация аминокислот. Поэтому на начальном этапе голодания мышцы теряют аминокислоты, которые идут на энергетические нужды. В дальнейшем организм адаптируется к отсутствию новых поступлений аминокислот (снижается потребность в зависящем от белка глюконеогенезе за счет использования энергетического потенциала кетоновых тел) и потеря белка мускулатуры уменьшается.
Читайте также: Евротекс мебельная ткань рогожка образцы название
Хотите больше новой информации по вопросам диетологии?
Оформите подписку на информационно-практический журнал «Практическая диетология»!
Роль почек
Почки не только выводят конечные продукты азотного распада (мочевину, креатинин и др.), но и являются дополнительным местом ресинтеза глюкозы из аминокислот, а также регулируют образование аммиака, компенсируя избыток ионов водорода в крови.
Глюконеогенез и функционирование кислотно-щелочной регуляции тесно скоординированы, поскольку субстраты этих процессов появляются при дезаминировании аминокислот: углерод для синтеза глюкозы и азот — для аммиака. Существует даже мнение, что именно производство глюкозы является основной реакцией почек на ацидоз, а образование аммиака происходит вторично.
Белок в нервной ткани
Для нервной ткани характерны более высокие концентрации аминокислот, чем в плазме. Это позволяет обеспечить мозг достаточным количеством ароматических аминокислот, являющихся предшественниками нейромедиаторов.
Некоторые заменимые аминокислоты, такие как глутамат (из которого при участии пиридоксина образуется гамма-аминомасляная кислота) и аспартат, также обладают влиянием на возбудимость нервной ткани. Их концентрация здесь высока, при этом заменимые аминокислоты способны синтезироваться и на месте.
Сон после еды
Специфическую роль играет триптофан, являющийся предшественником серотонина. Именно с повышением концентрации триптофана (а следовательно, и серотонина) связана сонливость после еды. Такой эффект особенно выражен при приеме больших количеств триптофана совместно с углеводистой пищей. Повышенная секреция инсулина снижает уровень в крови аминокислот с ветвящимися цепями, которые при преодолении барьера «кровь — мозг» обладают конкурентными взаимоотношениями с ароматическими аминокислотами, но в то же время не оказывает влияния на концентрацию связанного с альбумином триптофана. Благодаря подобным эффектам препараты триптофана могут использоваться в психиатрической практике.
При заболеваниях печени
Ограничение ароматических аминокислот в рационе, в связи с их влиянием на центральную нервную систему, имеет профилактическое значение при ведении пациентов с печеночной энцефалопатией. Элементные аминокислотные диеты с преимущественным содержанием лейцина, изолейцина, валина и аргинина помогают избежать развития белковой недостаточности у гепатологических больных и в то же время не приводят к возникновению печеночной комы.
Основные пластические функции протеиногенных аминокислот перечислены в таблице 2.
Таблица 2. Основные функции аминокислот
| Аланин | Предшественник глюконеогенеза, переносчик азота из периферических тканей в печень |
| Аргинин | Непосредственный предшественник мочевины |
| Аспарагиновая кислота | Предшественник глюконеогенеза, предшественник пиримидина, используется для синтеза мочевины |
| Глутаминовая кислота | Донор аминогрупп для многих реакций, переносчик азота (проникает через мембраны легче, чем глутамин), источник аммиака, предшественник ГАМК |
| Глицин | Предшественник пуринов, глютатиона и креатинина, входит в состав гемоглобина и цитохромов, нейротрансмиттер |
| Гистидин | Предшественник гистамина, донор углерода |
| Лизин | Предшественник карнитина (транспорт жирных кислот), составляющая коллагена |
| Метионин | Донор метальных групп для многих синтетических процессов (в т. ч. холина, пиримидинов), предшественник цистеина, участвует в метаболизме никотиновой кислоты и гистамина |
| Фенилаланин | Предшественник тирозина |
| Серин | Составляющая фосфолипидов, предшественник сфинголипидов, предшественник этаноламина и холина, участвует в синтезе пуринов и пиримидинов |
| Триптофан | Предшественник серотонина и никотинамида |
| Тирозин | Предшественник катехоламинов, допамина, меланина, тироксина |
| Цистеин | Предшественник таурина (желчные кислоты), входит в состав глютатиона (антиоксидантная система) |
Нормы потребления белка
Метаболизм белка в организме тесно связан с витамином В 6 , который является кофактором ферментов трансаминирования, обеспечивающих синтез заменимых и первый этап катаболизма всех аминокислот. Поэтому повышенный уровень потребления белка требует адекватного повышения количества пиридоксина.
Современные рекомендации по обеспечению пожилых людей и стариков основными питательными веществами, в первую очередь белками, свидетельствуют о целесообразном некотором снижении суточного количества белковых продуктов в пищевом рационе до 0,75–0,8 г/кг веса. Это связано с тем, что интенсивность основных физиологических функций с каждым десятилетием жизни человека после 50 лет снижается почти на 10 % (Rogers J., Jensen G., 2004), потребность белка уменьшается за счет инволюции синтетических и пластических процессов и ферментообразования, продукции гормонов, ряда биологически активных веществ, обеспечения мышечной деятельности и т. д.
Рекомендуемые нормы потребления для белка с учетом приведенных выше показателей составляют 55–62 г/сут (для мужчины весом 77 кг в возрасте 60–70 лет) и 45–52 г/сут (для женщины весом 65 кг в возрасте 60–70 лет) по выводам IV Американского национального исследования по оценке здоровья и питания (2006).
Вместе с тем установлено, что при сохранении физической активности пожилых людей (профессиональной физической нагрузки, занятий физкультурой, работы на дачном участке и т. п.) для поддержания азотного равновесия организма требуется повышение белкового обеспечения пожилого человека в количестве 1–1,25 г/кг в день. Эта же квота пищевого белка полностью обеспечит потребности пожилого человека, находящегося в состоянии стресса, болезни или ранения (Lowenthal D. T., 1990).
Рис. 1. Влияние пищевых веществ на развитие болезней избыточного питания (по А. А. Покровскому)

Дефицит белка = старение
Важно отметить, что организм пожилого человека очень чувствителен как к дефициту экзогенно поступающих белков, так и к их избытку. В условиях белкового дефицита прогрессирующе развиваются процессы дистрофии и атрофии клеточных структур, в первую очередь мышечной ткани, слизистых оболочек (желудочно-кишечного тракта, дыхательной системы и др.), паренхиматозных органов (поджелудочной железы, печени, эндокринных желез и др.), структур иммунной системы. Белковый дефицит питания активизирует процессы старения организма.
Механизмы патологического действия на организм пожилого и старого человека пищевой белковой перегрузки связаны в первую очередь с белковой «агрессией» печени и связанной с этим несостоятельностью ферментных систем, неполной деполимеризацией всех фракций белка, накоплением в крови токсических продуктов незавершенных окислительно-восстановительных реакций и т. д.
Белковая перегрузка
Интоксикационный процесс метаболического генеза при избыточном белковом питании пожилых и старых людей многократно усиливается по причине развития процессов гнилостной кишечной диспепсии в условиях относительной ферментной недостаточности желудка, поджелудочной железы, тонкой кишки и развития синдромов мальдигестии и мальабсорбции, а также кишечного дисбиоза (Барановский А. Ю., Кондрашина Э. А., 2008).
Белковая пищевая перегрузка в рамках интоксикационного синдрома способствует перевозбуждению центральной нервной системы, иногда — состояниям, близким к неврозам. При этом наблюдается повышенный расход витаминов в организме с формированием витаминной недостаточности.
При длительном высокобелковом питании вначале наблюдается компенсаторное усиление, а затем угнетение секреторной функции желудка и поджелудочной железы, повышается риск развития таких заболеваний, как подагра, мочекаменная болезнь.
В следующем выпуске журнала «Практическая диетология» мы продолжим рассказ о геронтологических особенностях основных видов обмена веществ пациентов пожилого и старческого возраста — углеводном и жировом обмене.
// ПД
Хотите больше новой информации по вопросам диетологии?
Оформите подписку на информационно-практический журнал «Практическая диетология»!
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом