
а) Фосфолипиды. Фосфолипиды в организме представлены в основном лецитином, кефалином и сфингомиелином. Типичная химическая структура этих веществ показана на рисунке ниже.
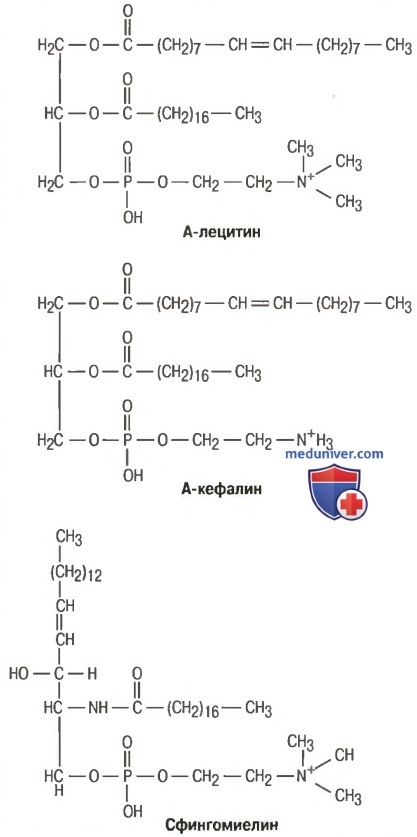
Типичные фосфолипиды
Фосфолипиды всегда содержат одну или несколько молекул жирных кислот, один остаток фосфорной кислоты и, кроме того, включают азотистое основание. Возможны небольшие вариации химической структуры фосфолипидов, но их физические свойства сходны, т.к. все они растворяются в липидах, транспортируются в виде липопротеинов и повсеместно используются в организме для пластических функций, образуя, например, наружные и внутриклеточные мембраны.
б) Образование фосфолипидов. Фосфолипиды синтезируются практически во всех клетках тела, хотя некоторые клетки обладают особыми возможностями для образования их в огромных количествах. Почти 90% фосфолипидов синтезируется в печени, существенные количества этих веществ образуются в эпителиоцитах кишечника во время всасывания жиров из кишки. Скорость образования фосфолипидов в ряде случаев определяется обычными факторами, в целом осуществляющими контроль за скоростью метаболизма жиров, поэтому, когда триглицериды депонируются в печени, скорость образования фосфолипидов увеличивается. Для образования некоторых фосфолипидов необходимы специфические вещества. Например, холин, присутствующий в рационе и синтезируемый в организме, необходим для образования лецитина, который, в свою очередь, нужен для формирования азотистого основания молекулы, или инозитол, необходимый для образования некоторых кефалинов.
в) Специфическое использование фосфолипидов. К функциям фосфолипидов относят следующие:
(1) фосфолипиды являются важной составляющей липопротеинов крови и необходимы для образования и функционирования большинства из них. При отсутствии фосфолипидов возникают серьезные нарушения транспорта холестерола и других фосфолипидов;
(2) тромбопластин, необходимый для инициации процесса свертывания, включает один из фосфолипидов как составную часть;
(3) большое количество сфингомиелинов присутствуют в нервной системе. По своим электрическим свойствам они являются изоляторами и присутствуют в окружающей нервные волокна миелиновой оболочке;
(4) фосфолипиды являются донорами фосфатных радикалов и используются в качестве таковых в химических реакциях в различных тканях;
(5) возможно, важнейшей функцией фосфолипидов является их участие в образовании структурных компонентов, главным образом мембран всех клеток организма, что обсуждается в отдельных статьях на сайте (просим вас пользоваться формой поиска выше) в связи с аналогичной функцией холестерола.
Физиология образования холестерола
а) Холестерол. Холестерол, формула которого приведена на рисунке ниже, присутствует в рационе всех людей и может медленно всасываться из желудочно-кишечного тракта в лимфатические сосуды кишечника.
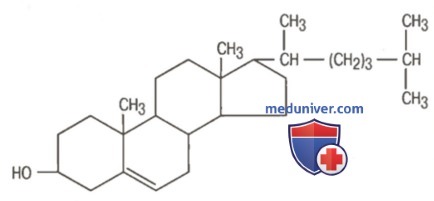
Холестерол
Он обладает высокой растворимостью в жирах и плохо растворим в воде. К специфическим свойствам холестерола относится его способность образовывать эфиры с жирными кислотами. Около 70% холестерола в липопротеинах плазмы крови представлено эфирами холестерола.
б) Образование холестерола. Холестерол, который абсорбируется в желудочно-кишечном тракте, называют экзогенным холестеролом. В большом количестве холестерол образуется в организме, в этом случае его называют эндогенным холестеролом. Эндогенный холестерол, циркулирующий в виде липопротеинов в плазме крови, образуется главным образом в печени, но практически все клетки могут образовывать какие-то его количества, т.к. фактически все мембраны структурных компонентов клеток частично образованы холестеролом.
Основным структурным компонентом холестерола является стерольное кольцо. Оно синтезируется из нескольких молекул ацетил-КоА. В свою очередь, стерольное кольцо может быть различными способами преобразовано в химические цепочки, формирующие:
(2) холиевую кислоту, служащую структурным основанием желчных кислот, образующихся в печени;
(3) важные стероидные гормоны, секретируемые корой надпочечников, яичниками, семенниками (об этих гормонах говорится в отдельных статьях на сайте — просим вас пользоваться формой поиска выше).
в) Факторы, которые влияют на концентрацию холестерола в плазме. Обратная связь, обеспечивающая регуляцию концентрации холестерола в организме. К важным факторам, влияющим на концентрацию холестерола в плазме крови, относят следующие.
Читайте также: Материалы для цветов из ткани своими руками
1. Увеличение количества холестерола в дневном рационе слегка повышает концентрацию холестерола в плазме крови. Употребление экзогенного холестерола, повышающее его концентрацию в плазме, ингибирует важный для синтеза эндогенного холестерола фермент 3-гидрокси-3-метилглютарил-КоА, что создает внутреннюю контролирующую обратную связь, препятствующую чрезмерному увеличению концентрации холестерола в плазме крови. В результате концентрация холестерола в плазме обычно не увеличивается и не уменьшается более чем на ±15% при изменении концентрации его в рационе, хотя могут быть индивидуальные значительные различия.
2. Высоконасыщенные жиры в рационе увеличивают концентрацию холестерола в крови на 15-25%. Это является результатом увеличения депонирования жиров в печени, приводящего к увеличению в клетках количества ацетил-КоА, из которого синтезируется холестерол, поэтому, чтобы понизить концентрацию холестерола в крови (что бывает даже более важно, чем поддержание низкого уровня холестерола в рационе), необходимо придерживаться диеты с низким содержанием насыщенных жиров.
3. Употребление жиров с более высоким содержанием ненасыщенных жирных кислот обычно приводит к легкому и даже умеренно выраженному снижению концентрации холестерола в крови. Механизм такого влияния неизвестен, хотя фактически это наблюдение положено в основу существующих диетологических рекомендаций.
4. Отсутствие инсулина или гормонов щитовидной железы увеличивает концентрацию холестерола в крови, в то время как избыток гормонов щитовидной железы приводит к снижению его концентрации. Это влияние, возможно, связано с увеличением степени активности специфических ферментов, ответственных за обмен липидов.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Синтез фосфолипидов в тканях
• Основные клеточные фосфолипиды синтезируются в мембране ЭПР, главным образом, со стороны цитозоля.
• Регуляция синтеза новых липидов контролируется локализацией ферментов, участвующих в этом процессе.
• Биосинтез холестерина регулируется протеолизом фактора транскрипции, который интегрирован в мембрану ЭПР.
Наряду с транслокацией и подготовкой белков к секреторному процессу, эндоплазматический ретикулум (ЭПР) функционирует как место первичного образования клеточных фосфолипидов. Эти молекулы синтезируются в мембране эндоплазматического ретикулума (ЭПР) и затем распределяются по отдельным мембранам и органеллам внутри клетки. К их числу относится плазматическая мембрана, митохондрии и органеллы секреторного пути.
При возникновении необходимости клетка должна обладать способностью увеличивать количество мембран. Наиболее очевидно это проявляется при удвоении плазматической мембраны и всех органелл, которое происходит в ходе клеточного цикла. Наряду с этим, при гиперфункции, отдельные органеллы могут увеличиваться в размерах. Например, при созревании предшественников В-лимфоцитов в плазматические клетки, секретирующие антитела, ЭПР сильно увеличивается, что соответствует потребностям усиленного транспорта белков по секреторному пути.
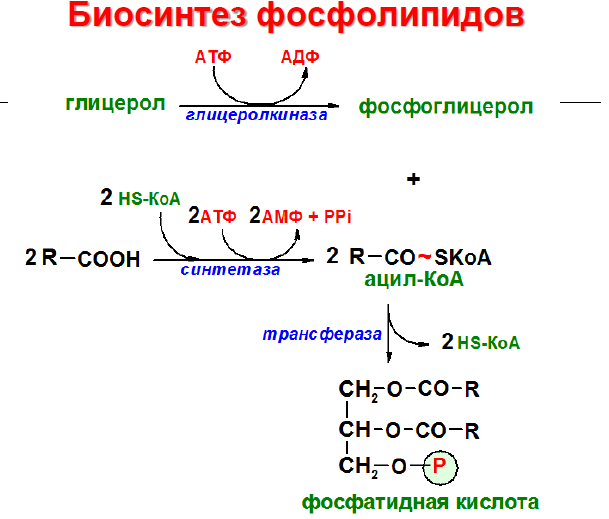
Образование фосфолипидов из растворимых предшественников de novo происходит, главным образом, на мембране ЭПР со стороны цитоплазмы, в ходе процесса, называемого цикл Кеннеди. Как показано на при росте мембраны ЭПР две молекулы жирных кислот, связанные с ацетил КоА, реагируют с глицерин-3-фосфатом, образуя диацилглицерин (ДАТ). В отличие от своих предшественников, ДАГ достаточно гидрофобен для того, чтобы интегрироваться в мембрану ЭПР.
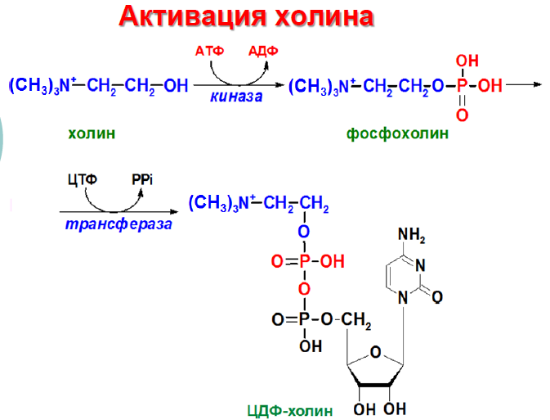
Попав в мембрану, ДАГ взаимодействует с «головой» будущей молекулы фосфолипида. Как показано на рисунке ниже, основные фосфолипиды различаются, главным образом, природой «головы» молекулы. Все они образуются при присоединении «головы» к ДАГ. Вначале происходит ее фосфорилирование, а затем присоединение к цитидиндифосфату (ЦДФ). После этого «голова» с одной из фосфатных групп переносится на ДАГ, и синтез завершается.
Присоединение головной группы холина к ЦЦФ с образованием фосфатидилхолина иллюстрирует использование клеткой субклеточной локализации белков для регуляции процессов биосинтеза. В этой реакции участвует фермент цитидилтрансфераза (ЦТ), и она контролирует скорость всего процесса синтеза фосфатидилхолина. В клетке ЦТ находится в двух различных формах: в неактивной, в цитозоле, и в активной, с цитозольной стороны мембраны ЭПР.
Читайте также: Плательная ткань ниагара что это такое
Механизм, который обеспечивает существование двух этих форм фермента, представляется не вполне ясным. Очевидно, однако, что условия внутри клетки, которые сигнализируют о необходимости усиления синтеза фосфатидилхолина, могут контролировать переход ЦТ из одной формы в другую. Например, увеличение в клетке концентрации свободного ДАТ или жирных кислот вызывает миграцию ЦТ на мембрану ЭПР, где эти компоненты реагируют с холином. В то же время, когда в клетке содержится много фосфатидилхолина, ЦТ локализуется в цитозоле. Таким образом, предотвращается синтез фосфатидилхолина.
Наряду с фосфатидилхолином, который является наиболее распространенным мембранным фосфолипидом, в цикле Кеннеди также образуются фосфатидилэтаноламин и фосфатидилинозитол. У дрожжей S. cerevisiae синтезируется фосфатидилсерин. У животных, однако, этот фосфолипид образуется за счет другой реакции, которая не требует участия ЦЦФ, однако также протекает с цитозольной стороны мембраны ЭПР.
Интересная особенность синтеза фосфатидилэтаноламина заключается в том, что хотя этот фосфолипид образуется в цикле Кеннеди, он также может синтезироваться в митохондриях. В этих органеллах он образуется при модификации фосфатидилсерина, который образовался в ЭПР. Образование фосфатидилэтаноламина в клеточных компартментах, отличающихся от тех, в которых синтезируется его предшественник, предполагает существование механизма транспорта липидов между двумя компартментами.
Имеются данные, позволяющие предполагать, что этот транспорт осуществляется посредством специального субдомена ЭПР, который называется мембраной, связанной с митохондрией (МСМ). МСМ представляет собой участок ЭПР, который физически ассоциирован с митохондрией. Он характеризуется плоской поверхностью и тесным контактом с органеллой. В МСМ находятся ферменты синтеза фосфатидилсерина. Возможно, что область контакта мембраны с митохондрией обеспечивает быстрый перенос фосфатидилсерина в органеллу, хотя механизм этого переноса остается неясным.
Наряду с участием в синтезе фосфолипидов, эндоплазматический ретикулум (ЭПР) является местом синтеза стеринов. К числу основных стеринов мембран клетки относится холестерин; его биосинтез осуществляется в несколько этапов. Первые несколько реакций происходят в цитозоле, а остальные в мембране ЭПР. Также в ЭПР находятся элементы, необходимые для регуляции образования холестерина. Ключевыми медиаторами этого регуляторного пути являются белки, связывающиеся с регуляторным стеринреспонсивным элементом (SREBP).
Эти белки обычно интегрированы в мембрану эндоплазматического ретикулума (ЭПР) и со стороны люмена содержат небольшую петлю, соединяющую два трансмембранных домена, N- и С-концевые участки которых ориентированы в сторону цитозоля. Также в ЭПР находится белок SCAP (белок, активирующий разрезание SREBP). Этот белок контролирует уровень внутриклеточного холестерина. Когда возникает необходимость в повышенном синтезе холестерина, SCAP транспортирует SREBP из ЭПР в аппарат Гольджи. Там происходит разрезание SREBP, и N-терминальный участок молекулы высвобождается в цитозоль. (В реакции участвуют те же протеазы, которые расщепляют АТФ 6 при возникновении стрессорных условий в ЭПР. Затем N-терминальный домен SREBP мигрирует в ядро, где действует как фактор транскрипции, активируя экспрессию генов в каскаде реакций биосинтеза холестерина.
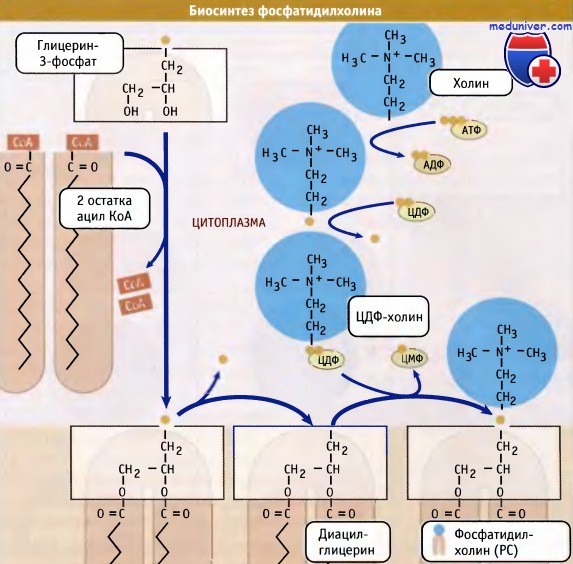 При объединении водорастворимых цитоплазматических компонентов (глицерин-3-фосфата, ацил КоА и полярной головки) образуется молекула фосфолипида.
При объединении водорастворимых цитоплазматических компонентов (глицерин-3-фосфата, ацил КоА и полярной головки) образуется молекула фосфолипида.
Новая растворимая в липидах молекула включается в мембрану. 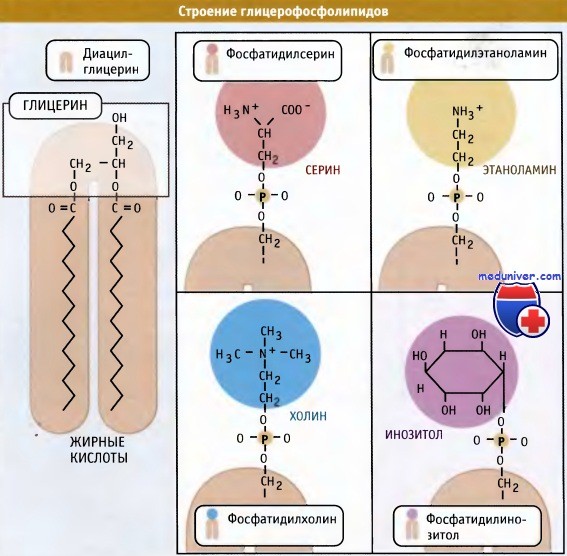 Основные мембранные фосфолипиды различаются по составу головок.
Основные мембранные фосфолипиды различаются по составу головок.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Биосинтез фосфолипидов в тканях, химизм, физиологическое значение.
Синтез фосфолипидов может происходить в различных тканях и по нескольким путям: в кишечнике с использованием пищевого холина или этаноламина (ресинтез фосфолипидов) и в печени (собственно биосинтез)
Читайте также: Халат из полиэстера что за ткань
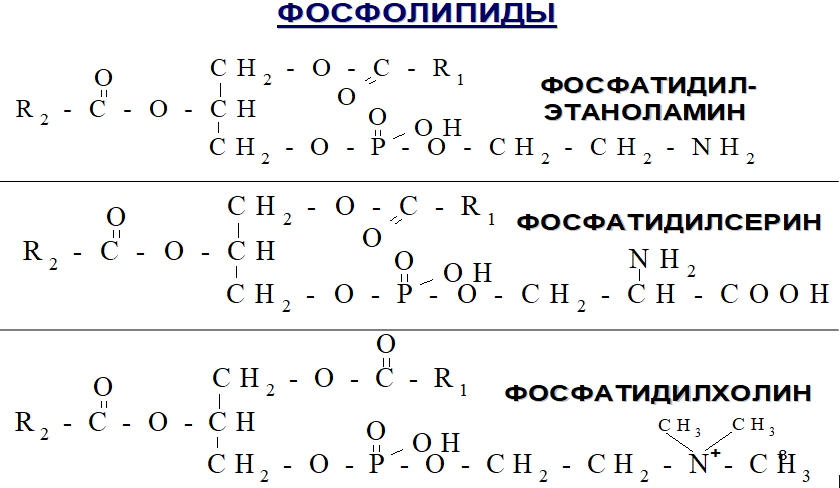
С интез фосфатидилхолинов, фосфатидилэтаноламинов и фосфатидилсеринов. Начальные этапы синтеза глицерофосфолипидов и жиров происходят одинаково до образования фосфатидной кислоты. Фосфатидная кислота может синтезироваться двумя разными путями: через глицеральдегид-3-фосфат и через дигидроксиацетонфосфат
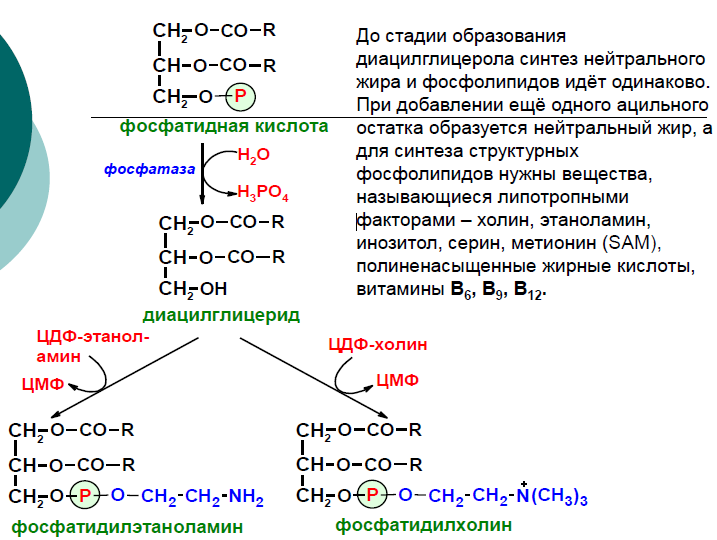
Н а следующем этапе фосфатидаза отщепляет от фосфатидной кислоты фосфатный остаток, в результате чего образуется диацилглицерол. Дальнейшие превращения диацилглицерола также могут идти разными путями. Один из вариантов — образование активной формы «полярной головки» фосфолипида: холин, серии или этаноламин превращаются в ЦДФ-холин, ЦДФ-серин или ЦДФ-этаноламин. Далее диацилглицерол взаимодействует с ЦМФ-производными, при этом выделяется ЦМФ, и образуется соответствующий фосфолигид, например, фосфатидилхолин. Между глицерофосфолипидами возможны различные взаимопревращения. Фосфатидилхолин может образовываться и другим путём: из фосфатидилэтаноламина, получая последовательно 3 метальные группы от SAM. Фосфатидилсерин может превращаться в фосфатидилэтаноламин путём декарбоксилирования. Фосфатидилэтаноламин может превращаться в фосфатидилсерин путём обмена этаноламина на серии.
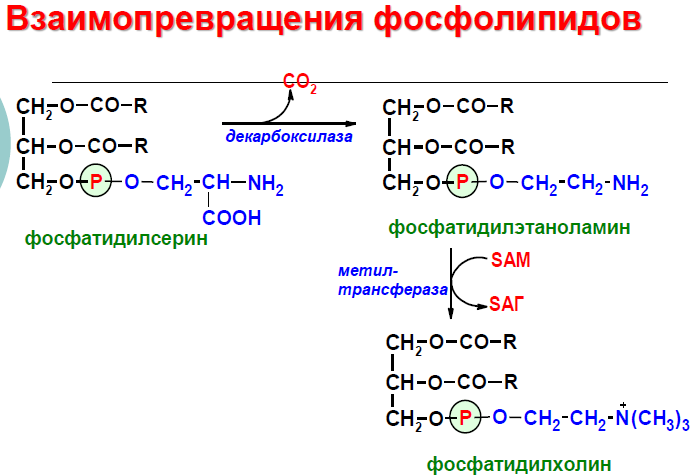
Б иологическая роль.
Входят в состав мембран, участвуя в их избирательной проницаемости.
Дипальмитоилфосфатидилхолин (до 80%), является компонентом сурфактанта, выстилающего альвеолы легких и препятствующей слипанию стенок альвеол во время вдоха.
Фосфатидилэтаноламин и фосфатидилхолин мембран взаимодействует с ферментами, образуя комплексы, которые активируют факторы свертывания крови. Фосфатидилсерин активирует процессы фибринолиза, связываясь с другими ферментами.
Обладают липотропным действием, препятствуя отложению нейтрального жира в печени.
Продукты их распада участвуют в патогенезе бронхиальной астмы, атеросклероза.
Фосфатидилсерин влияет на освобождение гистамина.
Холестерин, строение, биосинтез, судьба в организме.
Х 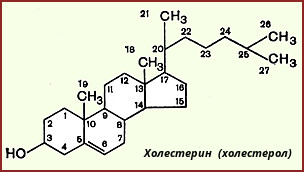
 олестерол — стероид, характерный только для животных организмов. Он синтезируется во многих тканях человека, но основное место синтеза — печень. В печени синтезируется более 50% холестерола, в тонком кишечнике — 15-20%, остальной холестерол синтезируется в коже, коре надпочечников, половых железах. В сутки в организме синтезируется около 1 г холестерола; с пищей поступает 300-500 мг. Холестерол выполняет много функций: входит в состав всех мембран клеток и влияет на их свойства, служит исходным субстратом в синтезе жёлчных кислот и стероидных гормонов. Предшественники в метаболическом пути синтеза холестерола превращаются также в убихинон — компонент дыхательной цепи и долихол, участвующий в синтезе гликопротеинов.
олестерол — стероид, характерный только для животных организмов. Он синтезируется во многих тканях человека, но основное место синтеза — печень. В печени синтезируется более 50% холестерола, в тонком кишечнике — 15-20%, остальной холестерол синтезируется в коже, коре надпочечников, половых железах. В сутки в организме синтезируется около 1 г холестерола; с пищей поступает 300-500 мг. Холестерол выполняет много функций: входит в состав всех мембран клеток и влияет на их свойства, служит исходным субстратом в синтезе жёлчных кислот и стероидных гормонов. Предшественники в метаболическом пути синтеза холестерола превращаются также в убихинон — компонент дыхательной цепи и долихол, участвующий в синтезе гликопротеинов.
1 стадия – синтез мевалоновой кислоты
Регуляция ключевого фермента синтеза хо-лестерола (ГМГ-КоА-редуктазы) происходит разными способами.
П 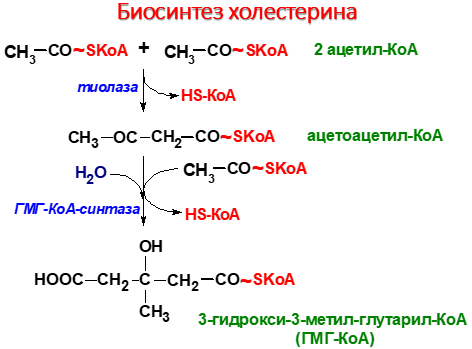
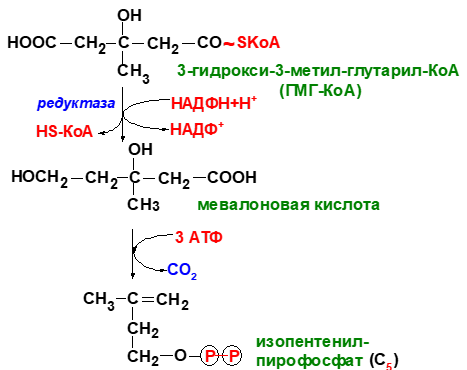 ри увеличении соотношения инсулин/глюкагон этот фермент дефосфорилируется и переходит в активное состояние. Действие инсулина осуществляется через 2 фермента: фосфатазу киназы ГМГ-КоА-редуктазы, которая превращает киназу в неактивное дефосфо-рилированное состояние; фосфатазу ГМГ-КоА-редуктазыпутём превращения её в дефосфорилированное активное состояние. Результатом этих реакций служит образование дефосфорилированной активной формы ГМГ-КоА-редуктазы. Следовательно, в абсорбтивный период синтез холестерола увеличивается. В этот период увеличивается и доступность исходного субстрата для синтеза холестерола — ацетил-КоА (в результате приёма пищи, содержащей углеводы, так как ацетил-КоА образуется в основном при распаде глюкозы). В постабсорбтивном состоянии глюкагон через протеинкиназу А стимулирует фосфо-рилирование ГМГ-КоА-редуктазы, переводя её в неактивное состояние. Это действие усиливается тем, что одновременно глюкагон стимулирует фосфорилирование и инактивацию фосфатазы ГМГ-КоА-редук-тазы и фосфорилирование киназы ГМГ-КоА-редуктазы, удерживая, таким образом, ГМГ-КоАредуктазу в фосфорилированном неактивном состоянии. В результате синтез холестерола в постабсорбтивном периоде и при голодании ингибируется. Ингибирование синтеза ГМГ-КоА-редуктазы. Конечный продукт метаболического пути (холестерол) снижает скорость транскрипции гена ГМГ-КоА-редуктазы, подавляя таким образом собственный синтез. В печени активно идёт синтез жёлчных кислот из холестерола, поэтому и жёлчные кислоты (как конечные продукты синтеза) подавляют активность гена ГМГ-КоА-редуктазы. Так как молекула ГМГ-КоА-редук-тазы существует около 3 ч после синтеза, то ингибирование синтеза этого фермента
ри увеличении соотношения инсулин/глюкагон этот фермент дефосфорилируется и переходит в активное состояние. Действие инсулина осуществляется через 2 фермента: фосфатазу киназы ГМГ-КоА-редуктазы, которая превращает киназу в неактивное дефосфо-рилированное состояние; фосфатазу ГМГ-КоА-редуктазыпутём превращения её в дефосфорилированное активное состояние. Результатом этих реакций служит образование дефосфорилированной активной формы ГМГ-КоА-редуктазы. Следовательно, в абсорбтивный период синтез холестерола увеличивается. В этот период увеличивается и доступность исходного субстрата для синтеза холестерола — ацетил-КоА (в результате приёма пищи, содержащей углеводы, так как ацетил-КоА образуется в основном при распаде глюкозы). В постабсорбтивном состоянии глюкагон через протеинкиназу А стимулирует фосфо-рилирование ГМГ-КоА-редуктазы, переводя её в неактивное состояние. Это действие усиливается тем, что одновременно глюкагон стимулирует фосфорилирование и инактивацию фосфатазы ГМГ-КоА-редук-тазы и фосфорилирование киназы ГМГ-КоА-редуктазы, удерживая, таким образом, ГМГ-КоАредуктазу в фосфорилированном неактивном состоянии. В результате синтез холестерола в постабсорбтивном периоде и при голодании ингибируется. Ингибирование синтеза ГМГ-КоА-редуктазы. Конечный продукт метаболического пути (холестерол) снижает скорость транскрипции гена ГМГ-КоА-редуктазы, подавляя таким образом собственный синтез. В печени активно идёт синтез жёлчных кислот из холестерола, поэтому и жёлчные кислоты (как конечные продукты синтеза) подавляют активность гена ГМГ-КоА-редуктазы. Так как молекула ГМГ-КоА-редук-тазы существует около 3 ч после синтеза, то ингибирование синтеза этого фермента
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности