
Было установлено, что гликоген может синтезироваться практически во всех органах и тканях. Однако наибольшая его концентрация обнаружена в печени (2-6%) и мышцах (0,5-2%). Поскольку мышечная масса организма человека велика, то большая часть гликогена организма содержится в мышцах. Глюкоза из крови легко проникает в клетки органов и тканей, проходя через биологические мембраны клеток. Как только глюкоза поступает в клетку, она метаболизируется в ней в результате первой химической реакции. фосфорилирование глюкозы происходит в присутствии АТФ и фермента — гексокиназы. Глюкоза превращается в глюкозо-6-фосфат . Этот эфир глюкозы теперь будет использоваться в анаболических и катаболических реакциях. Глюкоза из клетки может выйти только после реакции гидролиза при участии глюкозо-6-фосфатазы. Этот фермент есть в печени, почках, эпителии кишечника. В других органах, тканях его нет. Процесс биосинтеза гликогена протекает в 4 стадии:
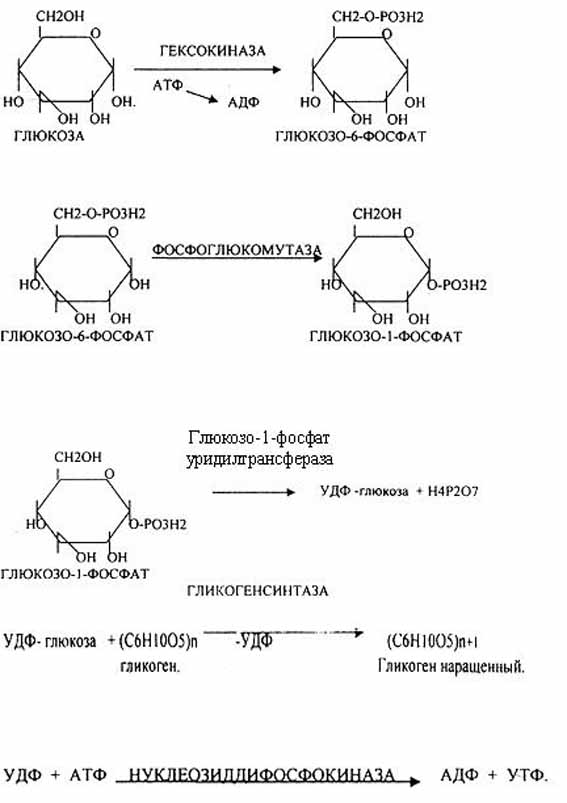
Гликогенсинтаза – является трансферазой, которая переносит остатки глюкозы, входящие в УДФ- глюкозу, на гликозидную связь остаточного в клетке гликогена, при этом образуется альфа(1,4)-гликозидные связи.
Образование альфа(1,6)-гликозидных связей в точках ветвления гликогена катализирует специальный гликоген-ветвящий фермент.
Образовавшийся в последней реакции УДФ, превращается в УТФ, при этом при биосинтезе гликогена поглощается еще 1 молекула АТФ.
Таким образом, на каждую молекулу глюкозы, включающуюся в структуру гликогена, расходуется 2 молекулы АТФ.
Гликоген в клетках накапливается во время пищеварения и рассматривается как резервная форма глюкозы, которая используется клетками в промежутках между приёмами пищи.
Существуют 2 пути распада гликогена в тканях:
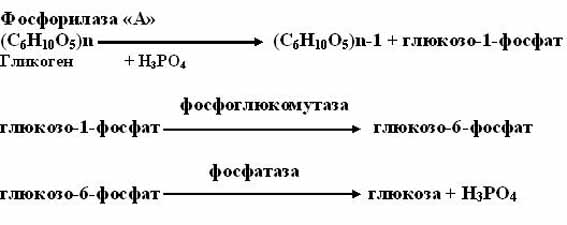
1. фосфоролитический путь (основной путь)
Протекает в печени, почках, эпителии кишечника. Схематически его можно записать в виде 3 реакций:
2. амилолитический путь (неосновной).
Протекает в печени при участии 3 ферментов: альфа -амилазы, амило-1,6-гликозидазы, гамма — амилазы.
Альфа – амилаза расщепляет в структуре крахмала альфа-1,4-гликозидные связи, амило-1,6-гликозидаза-гликозидные связи в точках ветвления, гамма-амилаза гидролизует концевые гликозидные связи в боковых ветвях гликогена.
ГЛИКОГЕНОЗЫ — болезни, связанные с нарушением процессов распада гликогена, при этом в клетках печени, почек, мышц гликоген накапливается в большом количестве. Клинически эти заболевания проявляются увеличением печени, мышечной слабостью, гипоглюкоземией натощак. Больные умирают в раннем детском возрасте. Наиболее часто встречаются следующие заболевания:
1.Болезнь Герса (генетический дефект фермента — фосфорилазы печени ).
2. Болезнь Мак-Ардля (генетический дефект фермента — фосфорилазы мышц ).
3. Болезнь Помпе (генетический дефект фермента — амило- 1,4-гликозидазы ).
4. Болезнь Кори (генетический дефект фермента — амило-1.6-гликозидазы ).
5. Болезнь Гирке (генетический дефект фермента — глюкозо-
АГЛИКОГЕНОЗЫ- болезни связанные с нарушением процессов синтеза гликогена в тканях. Характерными проявлениями нарушения синтеза гликогена являются: резкая гипогликемия натощак, рвоты, судороги, потеря сознания. Углеводное голодание клеток мозга приводит к нарушению психо-физического развития у детей. Смерть наступает в раннем детском возрасте. Наиболее часто встречаются:
1. Болезнь Льюиса (генетический дефект фермента – гликогенсинтазы ).
2. Болезнь Андерсена ( генетический дефект фермента — гликоген-ветвящего ).
Читайте также: Поролоновая подложка под ткань
Синтез гликогена (гликогеногенез)
Синтез гликогена (гликогеногенез)
Гликоген синтезируется в период пищеварения (через 1–2 часа после приема углеводный пищи). Синтез гликогена из глюкозы, как и любой анаболический процесс, является эндергоническим, т. е. требует затрат энергии.
Синтез гликогена включает 4 этапа:
1. Фосфорилирование глюкозы до глюкозо-6-фосфата при участии гексокиназы или глюкокиназы.
2. Активация первого углеродного атома с образованием активной формы – УДФ – глюкозы.
3. Образование ?-1,4-гликозидных связей. В присутствии «затравки» гликогена (молекулы, включающей не менее 4 остатков глюкозы) фермент гликогенсинтаза присоединяет остатки глюкозы из УДФ-глюкозы к С4-атому концевого остатка глюкозы в гликогене, образуя ?-1,4-гликозидную связь.
4. Образование ?-1,6-гликозидных связей (точки ветвления молекулы). Образование их осуществляется амилозо-1,4 ? 1,6-трансглюкозидазой (ветвящий или бранчинг фермент). Когда длина линейного участка цепи включает минимально 11 остатков глюкозы, этот фермент переносит фрагмент (1 ? 4) цепи с минимальным количеством 6 остатков глюкозы на соседнюю цепь или на несколько участков глюкозы дальше, образуя ?-1,6-гликозидную связь. Таким образом, образуется точка ветвления. Ветви растут путем последовательного присоединения (1–4)-глюкозильных единиц и дальнейшего ветвления.
Гликогенсинтаза – регуляторный фермент, существующий в двух формах:
1. – дефосфорилированной, активной (форма а);
2. – фосфорилированной, неактивной (форма b).
Активная форма образуется из неактивной под действием фосфатазы гликогенсинтазы при дефосфорилировании. Превращение активной формы в неактивную происходит при участии протеинкиназы путем фосфорилирования за счет АТФ.

Рис. 18.-1. Регуляция активности гликогенсинтазы.
Распад гликогена может проходить двумя путями.
1. Гидролитический – при участии амилазы с образованием декстринов и даже свободной глюкозы.
2. Фосфоролитический – под действием фосфорилазы и образованием глюкозо-1-фосфата. Это основной путь распада гликогена.
Фосфорилаза – сложный регуляторный фермент, существующий в двух формах – активной и неактивной. Активная форма (фосфорилаза а) – это тетрамер, в котором каждая субъединица соединена с остатком ортофосфата через гидроксильную группу серина. Под действием фосфатазы фосфорилазы происходит дефосфорилирование, отщепление 4 молекул фосфорной кислоты, и фосфорилаза а превращается в неактивную форму – фосфорилазу b, распадаясь на две димерные молекулы. Фосфорилаза b активируется путем фосфорилирования остатков серина за счет АТФ под действием фермента киназы фосфорилазы. В свою очередь этот фермент также существует в двух формах. Активная киназа фосфорилазы – фосфорилированный фермент, превращается в неактивную форму под действием фосфатазы. Активация киназы фосфорилазы осуществляется путем фосфорилирования за счет АТФ в присутствии ионов Mg 2+ протеинкиназой.
Регуляция синтеза и распада гликогена носит каскадный характер и происходит путем химической модификации ферментов.
Поскольку синтез и распад гликогена протекают по разным метаболическим путям, эти процессы могут контролироваться реципрокно. Влияние гормонов на синтез и распад гликогена осуществляется путем изменения в противоположных направлениях активности двух ключевых ферментов: гликогенсинтазы и гликогенфосфорилазы с помощью их фосфорилирования и дефосфорилирования. Инсулин стимулирует синтез гликогена и тормозит распад, адреналин и глюкагон обладают противоположным эффектом.
Данный текст является ознакомительным фрагментом.
Читайте также
5. Синтез рибосомных РНК
5. Синтез рибосомных РНК В обычных клетках синтез трех видов рРНК (28S, 18S и малой 5S) координирован, т. е. на одну молекулу 28S образуется одна молекула 18S и одна молекула 5S. Синтез 28S и 18S происходит в виде одного большого, общего для них предшественника (пре-рРНК), который затем
Читайте также: Ткань стеклокром п 3 0
6. Гормоны регулируют синтез желтка и белка
6. Гормоны регулируют синтез желтка и белка Мы уже говорили, что у позвоночных животных желток будущего яйца синтезируется в печени. Этот синтез стимулируется женскими половыми стероидными гормонами — эстрогенами (подробнее см. специальную главу). Один из таких гормонов
Великий синтез
Великий синтез Как связать эволюцию с генетикой. Можно ли подойти к вопросам изменчивости, борьбы за существование, отбора — словом, дарвинизма, исходя не из тех совершенно бесформенных, расплывчатых, неопределенных воззрений на наследственность, которые только и
2.4. Конфронтация или новый синтез?
2.4. Конфронтация или новый синтез? Наиболее оправданной позицией многим эволюционистам уже давно представлялся синтез положений СТЭ с концепциями направленной эволюции и сальтационизма на основе достижений генетики. Разные авторы говорили, что пора переходить от
3. ПОВТОРНЫЙ ВХОД ВОЗБУЖДЕНИЯ И ИНФОРМАЦИОННЫЙ СИНТЕЗ
3. ПОВТОРНЫЙ ВХОД ВОЗБУЖДЕНИЯ И ИНФОРМАЦИОННЫЙ СИНТЕЗ Описанная ранее концепция «светлого пятна» исходит из того, что сознание определяется некоторым уровнем возбудимости мозговых структур. Однако можно предположить, что этого недостаточно и в действительности
Глава 18. Обмен гликогена
Глава 18. Обмен гликогена Гликоген – основной резервный полисахарид в животных тканях. Он представляет собой разветвленный гомополимер глюкозы, в котором остатки глюкозы соединены в линейных участках ?-1,4-гликозидными связями, а в точках ветвления – ?-1,6- гликозидными
Нарушения обмена гликогена
Нарушения обмена гликогена Гликогеновые болезни – группа наследственных нарушений в основе которых лежит снижение или отсутствие активности ферментов, катализирующих реакции синтеза или распада гликогена. К данным нарушениям относятся гликогенозы и
Синтез жирных кислот
Синтез жирных кислот Синтез жирных кислот происходит в основном в печени, в меньшей степени – в жировой ткани и лактирующей молочной железе. Гликолиз и последующее окислительное декарбоксилирование пирувата способствуют увеличению концентрации ацетил-КоА в матриксе
5.5. Альтернативные теории и синтез идей эволюционизма
5.5. Альтернативные теории и синтез идей эволюционизма В рамках научной методологии эволюционизму нет альтернативы, поскольку такой альтернативой может служить только креационизм. Однако сам эволюционизм не является однородным течением. Хотя после чтения популярных
Передача гормонального сигнала: синтез, секреция, транспорт гормонов, их действие на клетки-мишени и инактивация
Передача гормонального сигнала: синтез, секреция, транспорт гормонов, их действие на клетки-мишени и инактивация В определении понятия «гормон» было указано несколько этапов распространения гормонального сигнала (рис. 2.6). Рис. 2.6. Этапы распространения гормонального
Синтез гликогена (гликогенез)
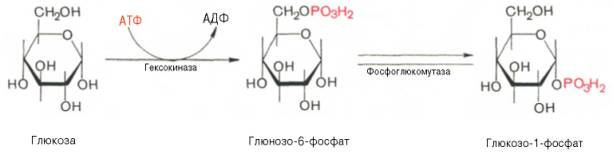
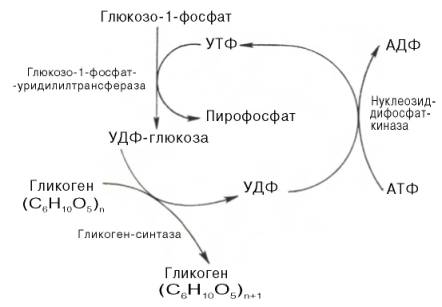
Прежде всего глюкоза подвергается фосфорилированию при участии фермента гексокиназы, а в печени – и глюкокиназы. Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фос-фат:
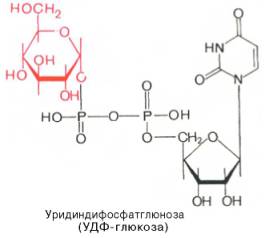
Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена. На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ (уридинтрифосфат), образуя уридиндифосфатглюкозу (УДФ-глюкоза) и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорилаза):
Читайте также: Дышащая ткань для одежды это
Глюкозо-1-фосфат + УТФ УДФ-глюкоза + Пирофосфат.
На второй стадии – стадии образования гликогена – происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную цепь гликогена («затравочное» количество). При этом образуется α-(1–>4)-связь между первым атомом углерода добавляемого остатка глюкозы и 4-гидроксильной группой остатка глюкозы цепи. Эта реакция катализируется ферментом гликогенсинтазой. Необходимо еще раз подчеркнуть, что реакция, катализируемая гликогенсинтазой, возможна только при условии, что полисахаридная цепь уже содержит более 4 остатков D-глю-козы.
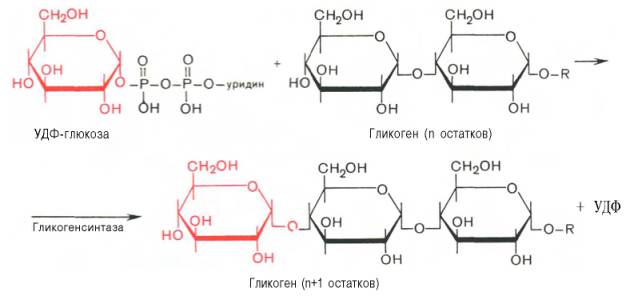
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и таким образом весь цикл превращений глюкозо-1-фосфата начинается сначала.
В целом образование α-1,4-глюкозидной ветви («амилозной» ветви) гликогена можно представить в виде следующей схемы:
Установлено, что гликогенсинтаза неспособна катализировать образование α-(1–>6)-связи, имеющейся в точках ветвления гликогена. Этот процесс катализирует специальный фермент, получивший название гли-когенветвящего фермента, или амило-(1–>4)–>(1–>6)-трансглюкозидазы. Последний катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6 или 7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксиль-ную группу остатка глюкозы той же или другой цепи гликогена. В результате образуется новая боковая цепь.
Ветвление повышает растворимость гликогена. Кроме того, благодаря ветвлению создается большое количество невосстанавливающих концевых остатков, которые являются местами действия гликогенфосфорилазы и гликогенсинтазы.
Таким образом, ветвление увеличивает скорость синтеза и расщепления гликогена.
Благодаря способности к отложению гликогена (главным образом в печени и мышцах и в меньшей степени в других органах и тканях) создаются условия для накопления в норме некоторого резерва углеводов. При повышении энерготрат в организме в результате возбуждения ЦНС обычно происходят усиление распада гликогена и образование глюкозы.
Помимо непосредственной передачи нервных импульсов к эффекторным органам и тканям, при возбуждении ЦНС повышаются функции ряда желез внутренней секреции (мозговое вещество надпочечников, щитовидная железа, гипофиз и др.), гормоны которых активируют распад гликогена, прежде всего в печени и мышцах (см. главу 8).
Как отмечалось, эффект катехоламинов в значительной мере опосредован действием цАМФ, который активирует протеинкиназы тканей. При участии последних происходит фосфорилирование ряда белков, в том числе гликогенсинтазы и фосфорилазы b – ферментов, участвующих в обмене углеводов. Фосфорилированный фермент гликогенсинтаза сам по себе малоактивен или полностью неактивен, но в значительной мере активируется положительным модулятором глюкозо-6-фосфатом, который увеличивает Vmaxфермента. Эта форма гликогенсинтазы называется D-формой, или зависимой (dependent) формой, поскольку ее активность зависит от глюкозо-6-фосфата. Дефосфорилированная форма гликоген-синтазы, называемая также I-формой, или независимой (independent) формой, активна и в отсутствие глюкозо-6-фосфата.
Таким образом, адреналин оказывает двойное действие на обмен углеводов: ингибирует синтез гликогена из УДФ-глюкозы, поскольку для проявления максимальной активности D-формы гликогенсинтазы нужны очень высокие концентрации глюкозо-6-фосфата, и ускоряет распад гликогена, так как способствует образованию активной фосфорилазы а. В целом суммарный результат действия адреналина состоит в ускорении превращения гликогена в глюкозу.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом