
Было установлено, что гликоген может синтезироваться практически во всех органах и тканях. Однако наибольшая его концентрация обнаружена в печени (2-6%) и мышцах (0,5-2%). Поскольку мышечная масса организма человека велика, то большая часть гликогена организма содержится в мышцах. Глюкоза из крови легко проникает в клетки органов и тканей, проходя через биологические мембраны клеток. Как только глюкоза поступает в клетку, она метаболизируется в ней в результате первой химической реакции. фосфорилирование глюкозы происходит в присутствии АТФ и фермента — гексокиназы. Глюкоза превращается в глюкозо-6-фосфат . Этот эфир глюкозы теперь будет использоваться в анаболических и катаболических реакциях. Глюкоза из клетки может выйти только после реакции гидролиза при участии глюкозо-6-фосфатазы. Этот фермент есть в печени, почках, эпителии кишечника. В других органах, тканях его нет. Процесс биосинтеза гликогена протекает в 4 стадии:
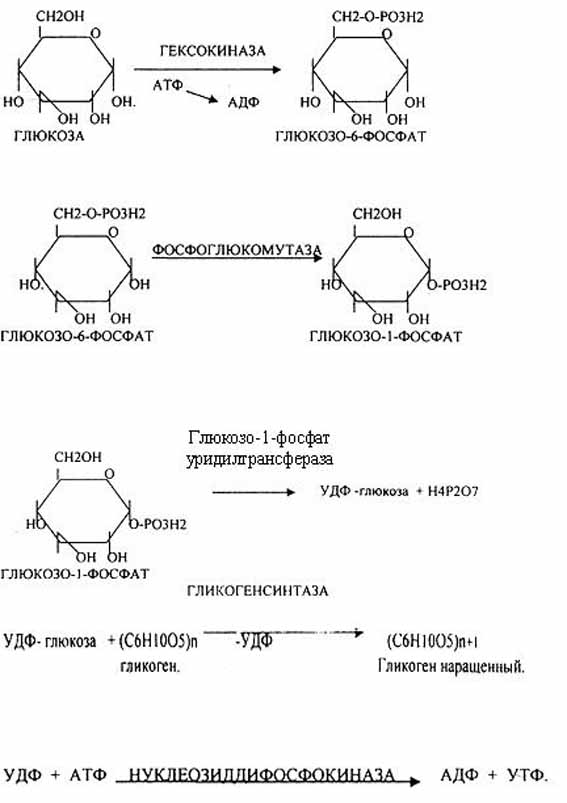
Гликогенсинтаза – является трансферазой, которая переносит остатки глюкозы, входящие в УДФ- глюкозу, на гликозидную связь остаточного в клетке гликогена, при этом образуется альфа(1,4)-гликозидные связи.
Образование альфа(1,6)-гликозидных связей в точках ветвления гликогена катализирует специальный гликоген-ветвящий фермент.
Образовавшийся в последней реакции УДФ, превращается в УТФ, при этом при биосинтезе гликогена поглощается еще 1 молекула АТФ.
Таким образом, на каждую молекулу глюкозы, включающуюся в структуру гликогена, расходуется 2 молекулы АТФ.
Гликоген в клетках накапливается во время пищеварения и рассматривается как резервная форма глюкозы, которая используется клетками в промежутках между приёмами пищи.
Существуют 2 пути распада гликогена в тканях:
1. фосфоролитический путь (основной путь)
Протекает в печени, почках, эпителии кишечника. Схематически его можно записать в виде 3 реакций:
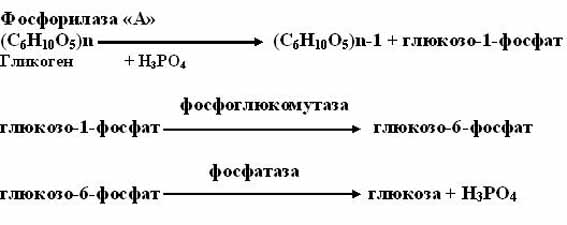
2. амилолитический путь (неосновной).
Протекает в печени при участии 3 ферментов: альфа -амилазы, амило-1,6-гликозидазы, гамма — амилазы.
Альфа – амилаза расщепляет в структуре крахмала альфа-1,4-гликозидные связи, амило-1,6-гликозидаза-гликозидные связи в точках ветвления, гамма-амилаза гидролизует концевые гликозидные связи в боковых ветвях гликогена.
ГЛИКОГЕНОЗЫ — болезни, связанные с нарушением процессов распада гликогена, при этом в клетках печени, почек, мышц гликоген накапливается в большом количестве. Клинически эти заболевания проявляются увеличением печени, мышечной слабостью, гипоглюкоземией натощак. Больные умирают в раннем детском возрасте. Наиболее часто встречаются следующие заболевания:
1.Болезнь Герса (генетический дефект фермента — фосфорилазы печени ).
2. Болезнь Мак-Ардля (генетический дефект фермента — фосфорилазы мышц ).
3. Болезнь Помпе (генетический дефект фермента — амило- 1,4-гликозидазы ).
4. Болезнь Кори (генетический дефект фермента — амило-1.6-гликозидазы ).
5. Болезнь Гирке (генетический дефект фермента — глюкозо-
АГЛИКОГЕНОЗЫ- болезни связанные с нарушением процессов синтеза гликогена в тканях. Характерными проявлениями нарушения синтеза гликогена являются: резкая гипогликемия натощак, рвоты, судороги, потеря сознания. Углеводное голодание клеток мозга приводит к нарушению психо-физического развития у детей. Смерть наступает в раннем детском возрасте. Наиболее часто встречаются:
Читайте также: Ткань чтобы сшить маску
1. Болезнь Льюиса (генетический дефект фермента – гликогенсинтазы ).
2. Болезнь Андерсена ( генетический дефект фермента — гликоген-ветвящего ).
7. Регуляция процессов синтеза и распада гликогена
Регуляция осуществляется на уровне двух ферментов: гликогенсинтетазы, участвующей в синтезе гликогена, ифосфорилазы, катализирующей расщепление гликогена.
Основным механизмом регуляции активности этих ферментов является их ковалентная модификация путем фосфорилирования-дефосфорилирования. Фосорилированная фосфорилаза или фосфорилаза «a» высокоактивна, в то же время фосфорилированная гликогенсинтетаза или синтетаза «b» неактивна. Таким образом, если оба фермента находятся в фосфорилированной форме, в клетке идет расщепление гликогена с образованием глюкозы. В дефосфорилированном состоянии, наоборот, неактивна фосфорилаза ( в форме «b») и активна гликогенсинтетаза ( в форме «a» ), в этой ситуации в клетке идет синтез гликогена из глюкозы.
Поскольку гликоген печени играет роль резерва глюкозы для всего организма, его синтез или распад должен контролироваться надклеточными регуляторными механизмами, работа которых должна быть направлена на поддержание постоянной концентрации глюкозы в крови. Эти механизмы должны обеспечивать включение синтеза гликогена в гепатоцитах при повышенных концентрациях глюкозы в крови и усиливать расщепление гликогена при падении содержания глюкозы в крови.
Итак, первичным сигналом, стимулирующим мобилизацию гликогена в печени, является снижение концентрации глюкозы в крови. В ответ на него альфа-клетки поджелудочной железы выбрасывают в кровь свой гормон — глюкагон. Глюкагон, циркулирующий в крови, взаимодействует со своим белком-рецептором, находящемся на внешней стороне наружной клеточной мембраны гепатоцита. образуя гор мон-рецепторный комплекс. Образование гормон-рецепторного комплекса приводит с помощью специального механизма к активации фермента аденилатциклазы, находящегося на внутренней поверхности наружной клеточной мембраны. Фермент катализирует образование в клетке циклической 3,5-АМФ ( цАМФ ) из АТФ. В свою очередь, цАМФ активирует в клетке фермент цАМФ-зависимую протеинкиназу. Неактивная форма протеинкиназы представляет собой олигомер, состоящий из четырех субъединиц: 2 регуляторных и двух каталитических. При повышении концентрации цАМФ в клетке к каждой из регуляторных субъединиц протеинкиназы присоединяется по 2 молекулы цАМФ, конформация регуляторных субъединиц изменяется и олигомер распадается на регуляторные и каталитичес кие субъединицы. Свободные каталитические субъединицы катализирует фосфорилирование в клетке ряда ферментов, в том числе фосфорилирование гликогенсинтетазы с переводом ее в неактивное состояние, выключая таким образом синтез гликогена . Одновременно идет фос форилирование киназы фосфорилазы, а этот фермент, активируясь при его фосфорилировании, в свою очередь катализирует фосфорилирование фосфорилазы с переводом его в активную форму, т.е. в форму «a». В результате активации фосфорилазы включается расщепление гликогена и гепатоциты начинают поставлять глюкозу в кровь.
Попутно отметим, что при стимуляции расщепления гликогена в печени катехоламинами в качестве главных посредников выступают b — рецепторы гепатоцитов, связывающие адреналин. При этом происходит повышение содержания ионов Са в клетках, где они стимулируют Са/кальмодулинчувствительную киназу фосфорилазы, которая в свою очередь активирует фосфорилазу путем её фосфорилирования.
Читайте также: Правильно заправить угол ткани
Повышение концентрации глюкозы в крови является внешним сигналом для гепатоцитов в отношении стимуляции синтеза гликогена и связывания таким образом излишней глюкозы из русла крови.
Срабатывает следующий механизм: при повышении концентрации глюкозы в крови возрастает и ее содержание в гепатоцитах. Повышение концентрации глюкозы в гепатоцитах, в свою очередь, достаточно сложным путем активирует в них фермент фосфопротеинфосфатазу, которая катализирует отщепление от фосфорилированных белков остатков фосфорной кислоты. Дефосфорилирование активной фосфорилазы переводит ее в неактивную форму, а дефосфорилирование неактивной гликогенсинтетазы активирует фермент. В результате система переходит в состояние, обеспечивающие синтез гликогена из глюкозы [8,10].
В снижении фосфорилазной активности в гепатоцитах определенную роль играет гормон b-клеток поджелудочной железы инсулин. Он выделяется b-клетками в ответ на повышение содержания глюкозы в крови. Его связывание с инсулиновыми рецепторами на поверхности гепатоцитов приводит к активации в клетках печени фермента фосфодиэстеразы, катализирующего превращение цАМФ в обычную АМФ, не обладающую способность стимулировать образование активной протеинкиназы. Этим путем прекращается нарабатывание в гепатоцитах активной фосфорилазы, что также имеет значение для ингибирования расщепления гликогена.
Вполне естественно, что механизмы регуляции синтеза и распада гликогена в клетках различных органов имеют свои особенности. В качестве примера можно указать, что в миоцитах покоящихся мышц или мышц, выполняющих небольшую по интенсивности работу, практически нет фосфорилазы «a», но расщепление гликогена все же идет [10]. Дело в том, что мышечная фосфорилаза, находящаяся в дефосфорилированном состоянии или в форме «b», является аллостерическим ферментом и активируется имеющимися в миоцитах АМФ и неорганическим фосфатом. Активированная таким образом фосфорилаза «b» обеспечивает скорость мобилизации гликогена, достаточную для выполнения умеренной физической работы. Однако при выполнении интенсивной работы, в особенности если нагрузка резко возрастает, этого уровня мобилизации гликогена становится недостаточно. В таком случае срабатывают надклеточные механизмы регуляции. В ответ на внезапно возникшую потребность в интенсивной мышечной деятельности в кровь поступает гормон адреналин из мозгового вещества надпочечников. Адреналин, связываясь с рецепторами на поверхности мышечных клеток, вызывает ответную реакцию миоцитов, близкую по своему механизму к только что описанной реакции гепатоцитов на глюкагон. В мышечных клетках появляется фосфорилаза «a» и инактивируется гликогенсинтетаза, а образовавшийся гл-6-ф используется как энергетическое «топливо», окислительный распад которого обеспечивает энергией мышечное сокращение [8].
Следует заметить, что высокие концентрации адреналина, наблюдающиеся в крови людей в условиях эмоционального стресса, ускоряют расщепление гликогена в печени, повышая тем самым содержание глюкозы в крови — защитная реакция, направленная на экстренную мобилизация энергетических ресурсов.
Читайте также: Груз для ткани при раскройке
37. Биосинтез и распад гликогена в тканях. Биологическая роль этих процессов. Гликогеновые болезни.
Гликоген может синтезироваться практически во всех органах и тканях. Большая часть гликогена содержится в мышцах.
Глюкоза из крови легко проникает в клетки органов и тканей, проходя через биологические мембраны клеток. Как только глюкоза заходит в клетку, она превращается в глюкозо-6-фосфат. Процесс биосинтеза протекает в 4 стадии:

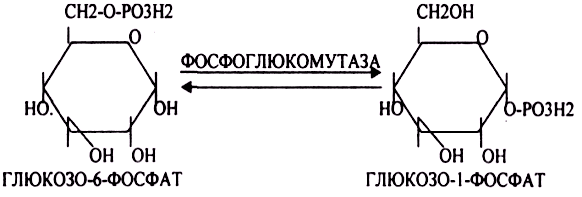
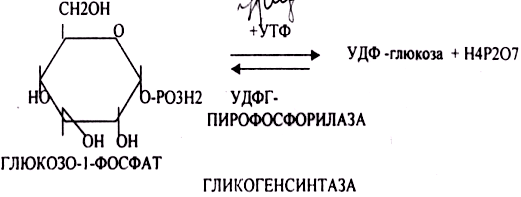

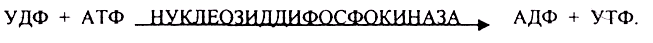
ГЛИКОГЕНСИНТАЗА — ТРАНСФЕРАЗА, которая переносит остатки глюкозы, входящие в УДФ- глюкозу, на ГЛИКОЗИДНУЮ связь остаточного в клетке гликогена. Образование альфа(1,6)-ГЛИКОЗИДНЫХ связей катализирует специальный ГЛИКОГЕН-ВЕТВЯЩИЙ фермент. Образовавшийся в последней реакции УДФ, превращается в УТФ, при этом поглощается I молекула АТФ. Гликоген в клетках накапливается во время пищеварения и рассматривается как резервная форма глюкозы, которая используется клетками в промежутках между приёмами пищи.
РАСПАД ГЛИКОГЕНА. Может идти 2 путями:
1. ОСНОВНОЙ ФОСФОРОЛИТИЧЕСКИЙ ПУТЬ
Он протекает в печени, почках, эпителии кишечника. Схематично можно записать в виде 3 реакций.

2.НЕОСНОВНОЙ АМИЛОЛИТИЧЕСКИЙ. Его доля незначительна. Протекает в печени при участии 3 ферментов: альфа — АМИЛАЗА, АМИЛО-1,6-ГЛИКОЗИДАЗА, гамма -АМИЛАЗА.
Это наследственное нарушение обмена гликогена, которое связано с недостатком какого-либо из ферментов, участвующих в синтезе или распаде гликогена. ГЛИКОГЕНОЗЫ — болезни, связанные с нарушением процессов распада гликогена. Клинически проявляются увеличением печени, мышечной слабостью, ГИПОГЛЮКОЗЭМИЕЙ натощак. Наиболее часто встречаются:
1.болезнь ФЕРСА — ФОСФОРИЛАЗА ПЕЧЕНИ.
2. Болезнь МАК-АРДЛЯ — ФОСФОРИЛАЗА МЫШЦ.
3. Болезнь ПОМПЕ- альфа- 1,4-ГЛИКОЗИДАЗА.
4. Болезнь КОРИ — АМИЛО-1.6-ГЛИКОЗИДАЗА.
5. Болезнь ГИРКЕ — ГЛЮКОЗО-6-ФОСФОТАЗА.
АГЛИКОГЕНОЗЫ характеризуются признаками нарушения синтеза. Клинически проявляются резкой ГИПОГЛЮКОЗЭМИЕЙ натощак, рвотой, судорогами, потерей сознания. Наиболее часто встречаются:
1. болезнь ЛЬЮИСА — ГЛИКОГЕНСИНТЕТАЗА.
2. Болезнь АНДЕРСЕНА — ГЛИКОГЕН-ВЕТВЯЩИЙ фермент.
38. Пути образования глюкозы в организме. Глюконеогенез. Возможные предшественники, последовательность реакций, биологическая роль.
ГЛЮКОНЕОГЕНЕЗ — это биосинтез глюкозы из неуглеводных предшественников, главными из которых являются ПИРУВАТ, ЛАКТАТ, ГЛИЦЕРИН, ряд АК, тем или иным путем превращающиеся в ФУМАРАТ, который в дальнейшем превращается в ЩУК. Другие АК превращаются в ПИРУВАТ. Главным местом синтеза глюкозы является печень. ГЛЮКОНЕОГЕНЕЗ обеспечивает синтез глюкозы, а также возврат ЛАКТАТА, образованного в реакциях анаэробного ГЛИКОЛИЗА, в клеточный фонд углеводов. За счет этого процесса поддерживается уровень глюкозы при углеводном голодании, сахарном диабете. Большинство реакций ГЛЮКОНЕОГЕНЕЗА представляют собой обратные реакции ГЛИКОЛИЗУ, за исключением трёх (ГЕКСОКИНАЗНОЙ, ФОСФОФРУКТОКИНАЗНОЙ, ПИРУВАТКИНАЗНОЙ), которые при ГЛюКОНЕОГЕНЕЗЕ имеют обходные пути.
Первая обходная реакция ГЛЮКОНЕОГЕНЕЗА связана с образованием 2-ФОСФОЕНОЛ ПВК и протекает в две стадии:

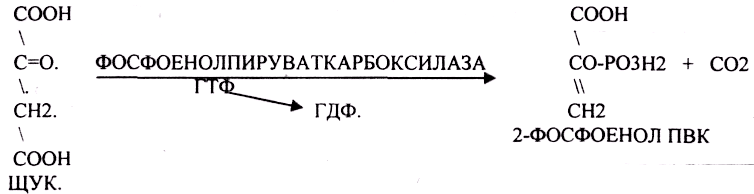
Вторая обходная реакция связана с образованием фруктозо-6-фосфата.
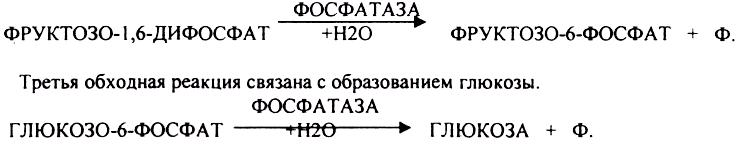
Образовавшаяся глюкоза может вновь использоваться клетками как пластический и энергетический материал, откладываться про запас в виде гликогена. В организме взрослого человека за сутки может быть образованно 80 гр. глюкозы.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом