
Установлено, что две наиболее распространенные мононасыщенные жирные кислоты — пальмитоолеиновая и олеиновая — синтезируются из пальмитиновой и стеариновой кислот. Двойная связь в молекулу указанных кислот вводится в микросомах клеток печени и жировой ткани при участии специфической оксигеназы и молекулярного кислорода. В этой реакции одна молекула кислорода используется в качестве акцептора двух пар электронов, одна пара из которых принадлежит субстрату (Ацил-КоА), а другая — НАДФН2:

Вместе с тем ткани человека и ряда животных неспособны синтезировать линолевую и линоленовую кислоты, а должны получать их с пищей (синтез этих кислот осуществляется растениями). В связи с этим линолевую и линоленовую кислоты, содержащие соответственно две и три двойные связи, называют незаменимыми жирными кислотами.
Все другие полиненасыщенные кислоты, обнаруженные у млекопитающих, образуются из четырех предшественников (пальмитоолеиноэой, олеиновой, линолевой и линоленовой киолот) путем дальнейшего удлинения цепи и (или) введения новых двойных связей. Происходит этот процесс при участии митохондриальных и микросомных ферментов. Например, синтез арахидоновой кислоты происходит по следующей схеме:

Биологическая роль полиненасыщенных жирных кислот в значительной мере прояснилась в связи с открытием нового класса физиологически активных соединений — простагландинов .
Есть основания считать, что скорость биосинтеза жирных кислот во многом определяется скоростью образования триглицеридов и фосфолипидов, ибо свободные жирные кислоты присутствуют в тканях и плазме крови в небольших количествах и в норме не накапливаются.
Синтез триглицеридов происходит из глицерина и жирных кислот (главным образом стеариновой, пальмитиновой и олеиновой). Путь биосинтеза триглицеридов в тканях протекает через образование глицерол-3-фосфата как промежуточного соединения. В почках, а также в стенке кишечника, где активность фермента глицеролкиназы высока, глицерин фосфорилируeтся АТФ с образованием глицерол-3-фосфата:

В жировой ткани и мышцах вследствие очень низкой активности глицеролкиназы образование глицерол-3-фосфата в основном связано с гликолизом или гликогенолизом 1 . 1 В тех случаях, когда содержание глюкозы в жировой ткани понижено (например, при голодании), образуется лишь незначительное количество глицерол-3-фосфата и освободившиеся в ходе липолиза свободные жирные кислоты не могут быть использованы на ресинтез триглицеридов, поэтому жирные кислоты покидают жировую ткань. Напротив, активация гликолиза в жировой ткани способствует накоплению в ней триглицеридов, а также входящих в их состав жирных кислот. Известно, что в процессе гликолитического распада глюкозы образуется диоксиацетонфосфат . Последний в присутствии цитоплазматической НАД-зависимой глицеролфосфатдегидрогеназы способен превращаться в глицерол-3-фосфат:

В печени же наблюдаются оба пути образования глицерол-3-фосфата.
Образовавшийся, тем или иным путем глицерол-3-фосфат ацилируется двумя молекулами КоА-производного жирной кислоты (т. е. «активными» формами жирной кислоты) 2 . 2 У некоторых микроорганизмов, например у Е. coli, донором ацильной группы являются не КоА-пронзводные, а АПБ-производные жирной кислоты. В результате образуется фосфатидная кислота:

Заметим, что хотя фосфатидная кислота и присутствует в клетках в чрезвычайно малых количествах, однако она является весьма важным промежуточным продуктом, общим для биосинтеза триглицеридов и глицерофосфолипидов (см. схему).

Если идет синтез триглицеридов, то происходит дефосфорилирование фосфатидной кислоты с помощью специфической фосфатазы (фосфатидатфосфатазы) и образование 1,2-диглицерида:

Биосинтез триглицеридов завершается этерификацией образовавшегося 1,2-диглицерида третьей молекулой ацил-КоА:

Синтез наиболее важных глицерофосфолипидов локализован главным образом в эндоплазматической сети клетки. Сначала фосфатидная кислота в результате обратимой реакции с цитидинтрифосфатом (ЦТФ) превращается в цитидиндифосфатдиглицерид (ЦДФ-диглицерид):

Затем в последующих реакциях, каждая из которых катализируется соответствующим ферментом, цитидинмонофосфат вытесняется из молекулы ЦДФ-диглицерида одним из двух соединений — серином или инозитом, образуя фосфатидилсерин или фосфатидилинозит, или 3-фосфатидил-глицерол-1-фосфат. В качестве примера приводим образование фосфатидилсерина:
Читайте также: Произведения искусства из ткани

В свою очередь фосфатидилсерин может декарбоксилироваться с образованием фосфатидилэтаноламина:

Фосфатидмлэтаноламин является предшественником фосфатидилхолина. В результате последовательного переноса трех метильных групп от трех молекул S-аденозилметионина (донора метальных групп) к аминогруппе остатка этаноламина образуется фосфатидилхолин:

Существует еще один путь синтеза фосфатидилэтаноламина и фосфатидилхолина в клетках животных. В этом пути также используется ЦТФ в качестве переносчика, но не фосфатидной кислоты, а фосфорилхолина или фосфорилэтаноламина (схема).


Еще в 60-х годах нынешнего столетия Блох и сотр. в опытах с использованием ацетата, меченного 14 С по метильной и карбоксильной группе, показал, что оба атома углерода уксусной кислоты включаются в холестерин печени приблизительно в одинаковых количествах. Кроме того, было доказано, что все атомы углерода холестерина происходят из ацетата.
В дальнейшем благодаря работам Линена, Редней, Поляка, Корнфорта, А. Н. Климова и других исследователей были выяснены основные детали ферментативного синтеза холестерина, насчитывающего более 35 энзиматических реакций. В синтезе холестерина можно выделить три основные стадии: первая — превращение активного ацетата в мевалоновую кислоту, вторая — образование сквалена из мевалоновой кислоты, третья — циклизация сквалена в холестерин.
Вначале рассмотрим стадию превращения активного ацетата в мевалоновую кислоту. Начальным этапом синтеза мевалоновой кислоты из ацетил-КоА является образование ацетоацетил-КоА посредством обратимой тиолазной реакции:

Затем последующая конденсация ацетоацетил-КоА с третьей молекулой ацетил-КоА при участии гидроксиметилглутарил-КоА-синтазы (ГМГ-КоА-синтазы) дает образование β-гидрокси-β-метилглутарил-КоА:

Заметим, что эти первые этапы синтеза мевалоновой кислоты нами уже рассматривались, когда речь шла об образовании кетоновых тел . Далее β-гидрокси-β-метилглутарил-КоА под влиянием НАДФ-зависимой гидроксиметилглутарил-КоА-редуктазы (ГМГ-КоА-редуктазы) в результате восстановления одной из карбоксильных групп и отщепления HS-KoA превращается в мевалоновую кислоту:

ГМГ-КоА-редуктазная реакция — первая практически необратимая реакция в цепи биосинтеза холестерина и протекает она со значителоной потерей свободной энергии (около 33,6 кДж). Установлено, что данная реакция лимитирует скорость биосинтеза холестерина.
Наряду с классическим путем биосинтеза мевалоновой кислоты имеется второй путь, в котором в качестве промежуточного субстрата образуется не β-гидрокси-β-метилглутарил-КоА, а β-гидрокси-β-метилглутарнл-S-АПБ. Реакции этого пути идентичны, по-видимому, начальным стадиям биосинтеза жирных кислот вплоть до образования ацетоацетил-S-АПБ. В образовании мевалоновой кислоты по этому пути принимает участие ацетил-КоА-карбоксилаза — фермент, осуществляющий превращение ацетил-КоА в малонил-КоА. Оптимальное соотношение малонил-КоА и ацетил-КоА для синтеза мевалоновой кислоты: две молекулы ацетил-КоА на одну молекулу малонил-КоА.
Участие малонил-КоА, основного субстрата биосинтеза жирных кислот, в образовании мевалоновой кислоты и различных полиизопреноидов показано для ряда биологических систем: печени голубя и крысы, молочной железы кролика, бесклеточных дрожжевых экстрактов. Этот путь биосинтеза мевалоновой кислоты отмечается преимущественно в цитоплазме клеток печени. Существенную роль в образовании мевалоната в данном случае играет гидроксиметилглутарил-КоА-редуктаза, обнаруженная в растворимой фракции печени крысы и неидентичная микросомному ферменту по ряду кинетических и регуляторных свойств. Известно, что микросомная гидроксиметилглутарил-КоА-редуктаза является основным звеном регуляции пути биосинтеза мевалоновой кислоты из ацетил-КоА с участием ацетоацетил-КоА-тиолазы и ГМГ-КоА-синтазы. Регуляция второго пути биосинтеза мевалоновой кислоты при ряде воздействий (голодание, кормление холестерином, введение поверхностно-активного вещества — тритона WR-1339) отличается от регуляции первого пути, в котором принимает участие микросомная редуктаза. Эти данные свидетельствуют о существовании двух автономных систем биосинтеза мевалоновой кислоты. Физиологическая роль второго пути изучена неокончательно. Полагают, что он имеет определенное значение не только для синтеза веществ нестероидной природы, таких, как боковая цепь убихинона и уникального основания N 6 (Δ 2 -изопентил)-аденозина некоторых тРНК, но и для биосинтеза стероидов (А. Н. Климов, Э. Д. Полякова).
Читайте также: Как сшить батон из ткани
Во второй стадии ситеза холестерина мевалоновая кислота превращается в сквален. Реакции второй стадии начинаются с фосфорилирования мевалоновой кислоты с помощью АТФ. В результате образуется 5′-пирофосфорный эфир, а затем 5′-пирофосфорный эфир мевалоновой кислоты:

5′-пирофосфомевалоновая кислота в результате последующего фосфорилирования третичной гидроксильной группы образует нестабильный промежуточный продукт — 3′-фосфо-5′-пирофосфомевалоновую кислоту, которая, декарбоксилируясь и теряя фосфорную кислоту, превращается в изопентенилпирофосфат. Последний изомеризуется в диметилаллилпирофосфат:

Затем эти два изомерных изопентенилпирофосфата (диметилаллилпирофосфат и изопентенилпирофосфат) конденсируются с высвобождением пирофосфата и образованием геранилпирофосфата. К геранилпирофосфату вновь присоединяется изопентенилпирофосфат, давая в результате этой реакции фарнезилпирофосфат:

В заключительной реакции данной стадии в результате восстановительной конденсации фарнезилпирофосфата с его диметилаллильным изомером образуется сквален:

В третьей стадии биосинтеза холестерина сквален под влиянием скваленоксидоциклазы циклизируется с образованием ланостерина:

Дальнейший процесс превращения ланостерина в холестерин включает ряд реакций, сопровождающихся удалением трех метальных групп, насыщением двойной связи в боковой цепи и перемещением двойной связи в кольце В из положения 8,9 в положение 5,6. Детали этих последних реакций во многом еще неизвестны:
Синтез липидов в тканях это
• Основные клеточные фосфолипиды синтезируются в мембране ЭПР, главным образом, со стороны цитозоля.
• Регуляция синтеза новых липидов контролируется локализацией ферментов, участвующих в этом процессе.
• Биосинтез холестерина регулируется протеолизом фактора транскрипции, который интегрирован в мембрану ЭПР.
Наряду с транслокацией и подготовкой белков к секреторному процессу, эндоплазматический ретикулум (ЭПР) функционирует как место первичного образования клеточных фосфолипидов. Эти молекулы синтезируются в мембране эндоплазматического ретикулума (ЭПР) и затем распределяются по отдельным мембранам и органеллам внутри клетки. К их числу относится плазматическая мембрана, митохондрии и органеллы секреторного пути.
При возникновении необходимости клетка должна обладать способностью увеличивать количество мембран. Наиболее очевидно это проявляется при удвоении плазматической мембраны и всех органелл, которое происходит в ходе клеточного цикла. Наряду с этим, при гиперфункции, отдельные органеллы могут увеличиваться в размерах. Например, при созревании предшественников В-лимфоцитов в плазматические клетки, секретирующие антитела, ЭПР сильно увеличивается, что соответствует потребностям усиленного транспорта белков по секреторному пути.
Образование фосфолипидов из растворимых предшественников de novo происходит, главным образом, на мембране ЭПР со стороны цитоплазмы, в ходе процесса, называемого цикл Кеннеди. Как показано на при росте мембраны ЭПР две молекулы жирных кислот, связанные с ацетил КоА, реагируют с глицерин-3-фосфатом, образуя диацилглицерин (ДАТ). В отличие от своих предшественников, ДАГ достаточно гидрофобен для того, чтобы интегрироваться в мембрану ЭПР.
Попав в мембрану, ДАГ взаимодействует с «головой» будущей молекулы фосфолипида. Как показано на рисунке ниже, основные фосфолипиды различаются, главным образом, природой «головы» молекулы. Все они образуются при присоединении «головы» к ДАГ. Вначале происходит ее фосфорилирование, а затем присоединение к цитидиндифосфату (ЦДФ). После этого «голова» с одной из фосфатных групп переносится на ДАГ, и синтез завершается.
Присоединение головной группы холина к ЦЦФ с образованием фосфатидилхолина иллюстрирует использование клеткой субклеточной локализации белков для регуляции процессов биосинтеза. В этой реакции участвует фермент цитидилтрансфераза (ЦТ), и она контролирует скорость всего процесса синтеза фосфатидилхолина. В клетке ЦТ находится в двух различных формах: в неактивной, в цитозоле, и в активной, с цитозольной стороны мембраны ЭПР.
Читайте также: Ткани от елены сорокиной
Механизм, который обеспечивает существование двух этих форм фермента, представляется не вполне ясным. Очевидно, однако, что условия внутри клетки, которые сигнализируют о необходимости усиления синтеза фосфатидилхолина, могут контролировать переход ЦТ из одной формы в другую. Например, увеличение в клетке концентрации свободного ДАТ или жирных кислот вызывает миграцию ЦТ на мембрану ЭПР, где эти компоненты реагируют с холином. В то же время, когда в клетке содержится много фосфатидилхолина, ЦТ локализуется в цитозоле. Таким образом, предотвращается синтез фосфатидилхолина.
Наряду с фосфатидилхолином, который является наиболее распространенным мембранным фосфолипидом, в цикле Кеннеди также образуются фосфатидилэтаноламин и фосфатидилинозитол. У дрожжей S. cerevisiae синтезируется фосфатидилсерин. У животных, однако, этот фосфолипид образуется за счет другой реакции, которая не требует участия ЦЦФ, однако также протекает с цитозольной стороны мембраны ЭПР.
Интересная особенность синтеза фосфатидилэтаноламина заключается в том, что хотя этот фосфолипид образуется в цикле Кеннеди, он также может синтезироваться в митохондриях. В этих органеллах он образуется при модификации фосфатидилсерина, который образовался в ЭПР. Образование фосфатидилэтаноламина в клеточных компартментах, отличающихся от тех, в которых синтезируется его предшественник, предполагает существование механизма транспорта липидов между двумя компартментами.
Имеются данные, позволяющие предполагать, что этот транспорт осуществляется посредством специального субдомена ЭПР, который называется мембраной, связанной с митохондрией (МСМ). МСМ представляет собой участок ЭПР, который физически ассоциирован с митохондрией. Он характеризуется плоской поверхностью и тесным контактом с органеллой. В МСМ находятся ферменты синтеза фосфатидилсерина. Возможно, что область контакта мембраны с митохондрией обеспечивает быстрый перенос фосфатидилсерина в органеллу, хотя механизм этого переноса остается неясным.
Наряду с участием в синтезе фосфолипидов, эндоплазматический ретикулум (ЭПР) является местом синтеза стеринов. К числу основных стеринов мембран клетки относится холестерин; его биосинтез осуществляется в несколько этапов. Первые несколько реакций происходят в цитозоле, а остальные в мембране ЭПР. Также в ЭПР находятся элементы, необходимые для регуляции образования холестерина. Ключевыми медиаторами этого регуляторного пути являются белки, связывающиеся с регуляторным стеринреспонсивным элементом (SREBP).
Эти белки обычно интегрированы в мембрану эндоплазматического ретикулума (ЭПР) и со стороны люмена содержат небольшую петлю, соединяющую два трансмембранных домена, N- и С-концевые участки которых ориентированы в сторону цитозоля. Также в ЭПР находится белок SCAP (белок, активирующий разрезание SREBP). Этот белок контролирует уровень внутриклеточного холестерина. Когда возникает необходимость в повышенном синтезе холестерина, SCAP транспортирует SREBP из ЭПР в аппарат Гольджи. Там происходит разрезание SREBP, и N-терминальный участок молекулы высвобождается в цитозоль. (В реакции участвуют те же протеазы, которые расщепляют АТФ 6 при возникновении стрессорных условий в ЭПР. Затем N-терминальный домен SREBP мигрирует в ядро, где действует как фактор транскрипции, активируя экспрессию генов в каскаде реакций биосинтеза холестерина.
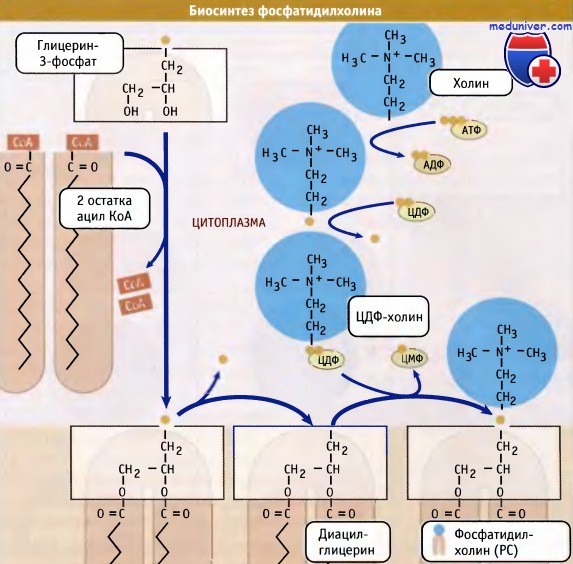 При объединении водорастворимых цитоплазматических компонентов (глицерин-3-фосфата, ацил КоА и полярной головки) образуется молекула фосфолипида.
При объединении водорастворимых цитоплазматических компонентов (глицерин-3-фосфата, ацил КоА и полярной головки) образуется молекула фосфолипида.
Новая растворимая в липидах молекула включается в мембрану. 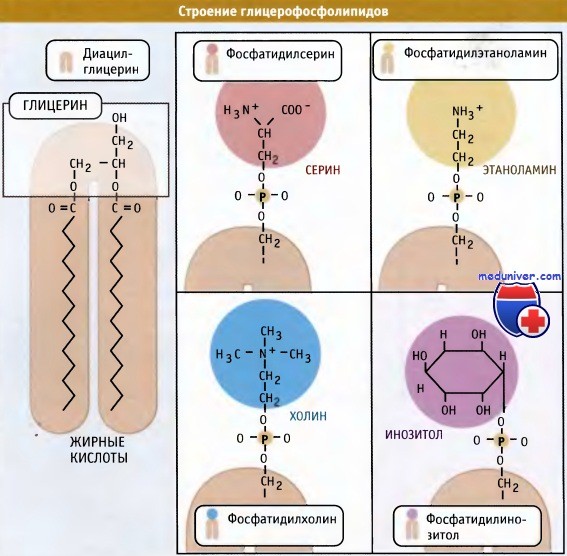 Основные мембранные фосфолипиды различаются по составу головок.
Основные мембранные фосфолипиды различаются по составу головок.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом