
Реакции биосинтеза липидов могут идти в гладкой эндоплазматической сети клеток всех органов. Субстратом для синтеза жиров de novo является глюкоза .
Как известно, попадая в клетку, глюкоза превращается в гликоген, пентозы и окисляется до пировиноградной кислоты. При высоком поступлении глюкоза используется для синтеза гликогена, но этот вариант ограничивается объемом клетки. Поэтому глюкоза «проваливается» в гликолиз и превращается в пируват либо напрямую, либо через пентозофосфатный шунт. Во втором случае образуется НАДФН, который понадобится впоследствии для синтеза жирных кислот.
Пируват переходит в митохондрии, декарбоксилируется в ацетил-SКоА и вступает в ЦТК . Однако в состоянии покоя, при отдыхе, при наличии избыточного количества энергии в клетке реакции ЦТК (в частности, изоцитратдегидрогеназная реакция) блокируются избытком АТФ и НАДН .
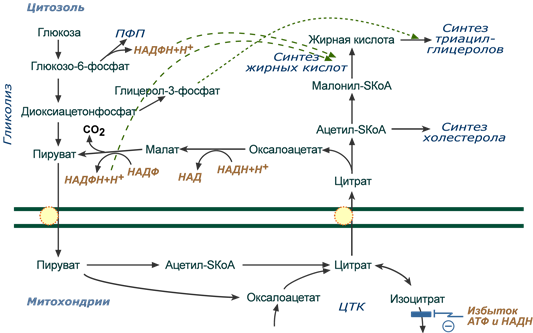
Общая схема биосинтеза триацилглицеролов и холестерола из глюкозы
В результате накапливается первый метаболит ЦТК – цитрат . По градиенту концентрации он перемещается в цитозоль, расщепляется с образованием ацетил-SКоА , который далее используется в биосинтезе холестерола и жирных кислот .
Оксалоацетат, также образуемый из цитрата, восстанавливается малатдегидрогеназой до яблочной кислоты и возвращается в митохондрии
- посредством малат-аспартатного челночного механизма (на рисунке не показан),
- после декарбоксилирования малата до пирувата НАДФ-зависимым малик-ферментом . Образованный НАДФН будет использован при синтезе жирных кислот или холестерина.
Липогенез.
АТФ необходим для синтеза активных форм субстратов, используемых в процессе синтеза жира.
Для синтеза нейтрального жира необходим глицерин в активной форме — глицерол-3-фосфат (фосфоглицерин). Глицерол-3-фосфат может быть получен двумя способами:
Путем активации глицерина с помощью глицеринкиназы.

2. Путем восстановления фосфодиоксиацетона, полученного при распаде глюкозы.
Кроме глицерина, для синтеза нейтрального жира необходимы жирные кислоты в активной форме. Активная форма любой жирной кислоты – Ацил-КоА. Образуется при участии фермента ацил-КоА-синтазы.

Здесь наблюдается глубокий распад АТФ до АМФ. АМФ не может вступить в окислительное фосфорилирование. Поэтому существует реакция: АТФ + АМФ 2АДФ. Поэтому затраты на активацию молекулы жирной кислоты эквивалентны затрате двух АТФ. Следующим этапом на пути синтеза жира является реакция образования фосфатидной кислоты:

Реакция катализируется ключевым ферментом липогенеза – глицерол-3-фосфатацилтрансферазой. Для этого фермента нет аллостерических эффекторов, но обнаружен адипсин (ацилстимулирующий белок), который облегчает взаимодействие Ацил-КоА с ферментом. Адипсин является продуктом протеолиза одного из компонентов системы комплемента. Относится к гормонам местного действия, так как вырабатывается в жировой ткани и действует там же.
Две последующие реакции являются завершающими в синтезе триацилглицерина.
Реакции синтеза не зависят от того, каково происхождение веществ – участников реакций.
Жир может синтезироваться как из продуктов распада жира, так и из углеводов. Синтез эндогенного жира из углеводов протекает в печени и в жировой ткани. Ацил-КоА синтезируется из Ацетил-КоА. ГБФ-путь распада углеводов обеспечивает синтез энергией. Образование Ацетил-КоА происходит в матриксе митохондрий. Синтез жирных кислот идет в цитоплазме на мембранах эндоплазматического ретикулума путем постепенного удлинения ацетил-КоА на 2 углеродных атома за каждый цикл. Удлинение высших жирных кислот, содержащих более 16 углеродных атомов, идёт путём реакций, обратных -окислению (О -окислении смотрите ниже, раздел «Катаболизм жирных кислот»). Однако реакции синтеза жирных кислот до 16 углеродных атомов принципиально отличаются от реакций, обратных -окислению. Они протекают обходным обратным путём.
Отличия реакций синтеза высших жирных кислот от обратных бета-окислению:
1. -окисление протекает в митохондриях, а синтез жирных кислот протекает в цитоплазме на мембранах эндоплазматического ретикулума. Но образуется Ацетил-КоА в митохондриях, а через мембраны сам проходить не может. Поэтому существуют механизмы транспорта Ацетил-КоА из митохондрий в цитоплазму.
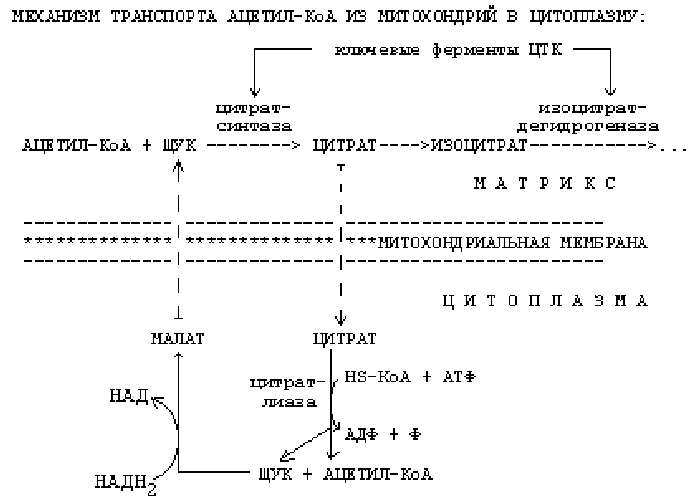
Ключевыми ферментами ЦТК являются цитратсинтаза и изоцитратдегидрогеназа. Основные аллостерические регуляторы этих ферментов — это АТФ и АДФ. Если в клетке много АТФ, то АТФ выступает как ингибитор этих ключевых ферментов. Однако изоцитратдегидрогеназа угнетается АТФ сильнее, чем цитратсинтетаза. Это приводит к накоплению цитрата и изоцитрата в матриксе митохондрии. При накоплении цитрат выходит из митохондрии в цитоплазму. В цитоплазме есть фермент цитратлиаза. Этот фермент расщепляет цитрат на ЩУК и Ацетил-КоА.
Условием для выхода Ацетил-КоА из митохондрии в цитоплазму является хорошее обеспечение клетки АТФ. Если АТФ в клетке мало, то Ацетил-КоА расщепляется до СО2 и Н2О.
2. В ходе -окисления промежуточные продукты связаны с HS-КоА, а при синтезе жирных кислот промежуточные продукты связаны с особым ацил-переносящим белком (АПБ). Это сложный белок. Его небелковая часть похожа по строению на КоА. Небелковая часть АПБ состоит из тиоэтиламина, пантотеновой кислоты (витамин В3) и фосфата.
Читайте также: У взрослых ген остеопротегерина экспрессирован во многих тканях
3. При -окислении в качестве окислителя используются НАД и ФАД. При синтезе ЖК нужен восстановитель — используется НАДФ*Н2.
В клетке существует 2 основных источника НАДФ*Н2 для синтеза жирных кислот:
а) ГМФ-путь распада углеводов
б) В реакции окисления малата (смотрите рисунок).
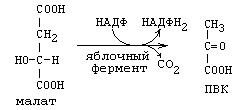
Эта реакция протекает в цитоплазме и катализируется ферментом МАЛАТДЕГИДРОГЕНАЗОЙ ДЕКАРБОКСИЛИРУЮЩЕЙ, тривиальное название которого – ЯБЛОЧНЫЙ ФЕРМЕНТ.
4. В ходе синтеза жирных кислот в каждом цикле удлинения используется не сам Ацетил-КоА, а его производное — малонил-КоА (при -окислении каждый цикл укорочения приводит к образованию Ацетил-КоА).
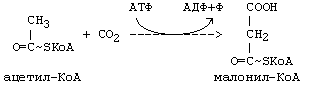
Эту реакцию катализирует фермент АЦЕТИЛ-КоА-КАРБОКСИЛАЗА. Это ключевой фермент в мультиферментной системе синтеза ЖК. Этот фермент регулируется по типу отрицательной обратной связи. Ингибитором является продукт синтеза: ацил-КоА с длинной цепью (n=16) — пальмитоил-КоА. Активатором является цитрат. В состав небелковой части этого фермента входит витамин H (биотин).
Далее происходит поэтапное удлинение молекулы Ац-КоА на 2 углеродных атома за каждый этап за счет малонил-КоА. В процессе удлинения малонил-КоА теряет СО2. После образования малонил-КоА основные реакции синтеза жирных кислот катализируются одним ферментом — синтетазой жирных кислот (фиксирован на мембранах эндоплазматического ретикулума). Синтетаза жирных кислот содержит 7 активных центров. Участок, связывающий малонил-КоА, содержит небелковый компонент – витамин B3 (пантотеновую кислоту).
РЕАКЦИИ СИНТЕЗА ЖИРНЫХ КИСЛОТ.
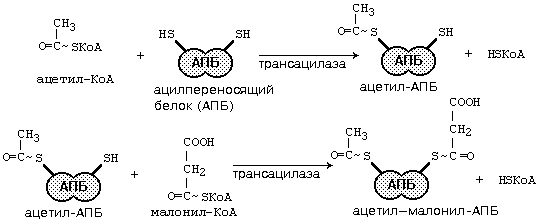
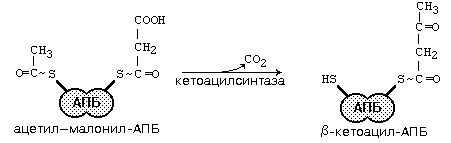
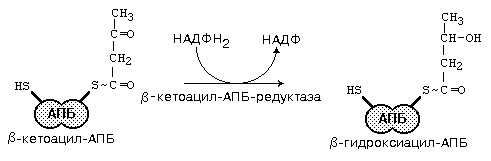
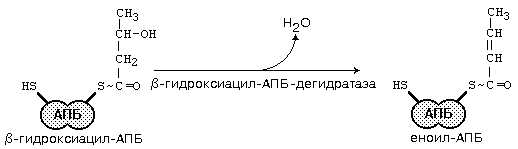

После этого ацил-АПБ вступает в новый цикл синтеза. К свободной SH-группе ацилпереносящего белка присоединяется новая молекула малонил-КоА. Затем происходит отщепление ацильного остатка, и он переносится на малонильный остаток с одновременным декарбоксилированием, и цикл реакций повторяется.
Таким образом, углеводородная цепочка будущей жирной кислоты постепенно растет (за каждый цикл – на два углеродных атома). Это происходит до момента, пока она не удлинится до 16 углеродных атомов (в случае синтеза пальмитиновой кислоты) или более (синтез других жирных кислот). Вслед за этим происходит тиолиз, и образуется в готовом виде активная форма жирной кислоты – ацил-КоА.
УСЛОВИЯ ДЛЯ ПРОТЕКАНИЯ СИНТЕЗА ВЫСШИХ ЖИРНЫХ КИСЛОТ:
1) Поступление углеводов, при окислении которых образуются необходимые субстраты и НАДФН2.
2) Высокий энергетический заряд клетки – высокое содержание АТФ, которое обеспечивает выход цитрата из митохондрий в цитоплазму.
Обмен углеводов и обмен жиров очень тесно связаны. Углеводы легко могут превращаться в жиры, а вот превращение жиров в углеводы невозможно. Жиры не могут превращаться в углеводы, так как Ацетил-КоА не может превращаться в пируват. Обмен жиров и углеводов объединяется как энергетический обмен, который находится под контролем гормонов.
Синтез нейтрального жира в тканях биохимия
а) Синтез триглицеридов из углеводов. Если когда-нибудь большие количества углеводов попадают в организм, они либо сразу используются для получения энергии, либо запасаются в виде гликогена, а избыток их быстро превращается в триглицериды и хранится в таком виде в жировой ткани. У человека большая часть триглицеридов образуется в печени, но очень небольшие количества могут образовываться и в самой жировой ткани. Триглицериды, образуемые в печени, транспортируются главным образом в виде липопротеинов очень низкой плотности в жировую ткань, где и хранятся.
б) Превращение ацетил-КоА в жирные кислоты. Первым этапом синтеза триглицеридов является превращение углеводов в ацетил-КоА.
Это происходит во время обычного расщепления глюкозы гликолитической системой. Вследствие того, что жирные кислоты являются крупными полимерами уксусной кислоты, легко представить, каким образом ацетил-КоА может быть превращен в жирную кислоту. Однако синтез жирных кислот не обеспечивается просто изменением направления реакции окислительного расщепления. Этот синтез осуществляется двуступенчатым процессом, показанным на рисунке ниже, с использованием малонил-КоА и НАДФ-Н в качестве основных посредников процесса полимеризации.
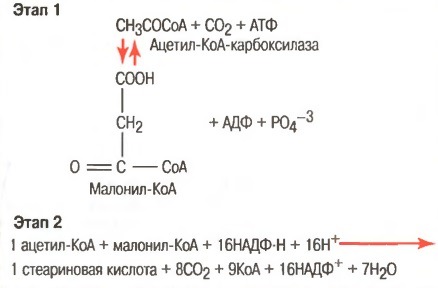
Синтез жирных кислот
в) Объединение жирных кислот с α-глицерофосфатом при образовании триглицеридов. Как только синтезируемые цепочки жирных кислот начинают включать от 14 до 18 атомов углерода, они взаимодействуют с глицеролом, образуя триглицериды. Ферменты, катализирующие эту реакцию, высокоспецифичны для жирных кислот с длиной цепочки от 14 атомов углерода и выше, что является фактором, контролирующим структурное соответствие триглицеридов, хранящихся в организме.
Как показано на рисунке ниже, образование глицероловой части молекулы триглицерида обеспечивается α-глицерофосфатом, который является побочным продуктом реакции гликолитического расщепления глюкозы.
Читайте также: Хорошо ли тянется ткань эластан
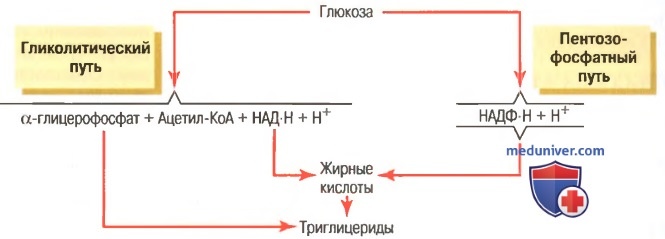
Общая схема синтеза триглицеридов из глюкозы
г) Эффективность превращения углеводов в жиры. Во время синтеза триглицеридов только 15% потенциально содержащейся в глюкозе энергии теряется в виде тепла. Остальные 85% преобразуются в энергию запасаемых триглицеридов.
д) Важность синтеза и хранения жиров. Синтез жиров из углеводов особенно важен в связи с двумя обстоятельствами.
1. Способность различных клеток организма запасать углеводы в виде гликогена выражена слабо. Только несколько сотен граммов гликогена может запасаться в печени, скелетных мышцах и всех других тканях организма, вместе взятых. В то же время могут запасаться килограммы жира, поэтому синтез жиров является способом, с помощью которого энергия, содержащаяся в избыточном количестве поступивших в организм углеводов (и белков), может запасаться, чтобы использоваться позднее. Количество энергии, которую запасает организм человека в виде жиров, приблизительно в 150 раз превышает количество энергии, запасаемой в виде углеводов.
2. Каждый грамм жиров содержит почти в 2,5 раза больше энергии, чем каждый грамм углеводов. Следовательно, при одном и той же массе тела организм может запасать в несколько раз больше энергии в виде жиров, чем в виде углеводов, что особенно важно, если необходима высокая степень подвижности, чтобы выжить.
е) Снижение синтеза жиров из углеводов при отсутствии инсулина. При отсутствии инсулина, как это бывает при тяжелом сахарном диабете, жиров синтезируется мало, если они вообще синтезируются, по следующим причинам. Во-первых, при отсутствии инсулина глюкоза не может попадать в сколько-нибудь существенных количествах в жировые ткани и клетки печени, что не обеспечивает образования достаточных количеств ацетил-КоА и НАДФ-Н, необходимых для синтеза жиров и получаемых при метаболизме глюкозы. Во-вторых, отсутствие глюкозы в жировых клетках существенно снижает количество наличного глицерофосфата, что также затрудняет образование триглицеридов.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Биосинтез нейтральных жиров и фосфолипидов
ОБМЕН ЛИПИДОВ
В тканях организма происходит непрерывное обновление липидов. Основную массу липидов тела человека составляют триглицериды, которыми особенно богата жировая ткань. В небольших количествах липиды присутствуют во всех тканях и органах. Поскольку липиды выполняют энергетическую функцию, то их метаболизм связан с процессами запасания и расхода энергии. Обмен фосфолипидов в первую очередь связан с процессами обновления биологических мембран, и естественно, этот процесс имеет место во всех органах и тканях.
Жиры являются необходимым компонентом пищевого рациона человека. В двенадцатиперстной кишке под действием желчи происходит эмульгирование жиров и их расщепление на глицерин и жирные кислоты под действием пищеварительных ферментов липаз. Продукты распада жиров всасываются в лимфу. Суточная потребность в жирах примерно составляет 100 г.
Катаболизм липидов
Первым этапом катаболизма липидов является их ферментативный гидролиз, который носит название липолиз. В результате этого процесса из нейтральных жиров образуется глицерин и три молекулы жирных кислот. При гидролизе фосфолипидов образуется глицерин, два остатка жирных кислот, остаток фосфорной кислоты и остаток радикала, который был связан с фосфорной кислотой, отличающийся у различных фосфолипидов (рис. 15).
Продукты гидролиза триглицеридов и фосфолипидов подвергаются дальнейшим метаболическим превращениям.
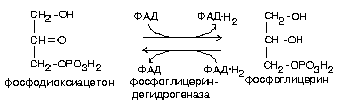
Обмен глицерина может осуществляться несколькими путями. Значительная часть глицерина, образовавшегося при гидролизе липидов, используется для их ресинтеза. Кроме того, продукты, образующиеся при окислении глицерина могут включаться в гликолиз или в глюконеогенез. В любом случае сначала происходит фосфорилирование глицерина с образованием глицерофосфата, донором фосфатной группы является молекула АТФ.
Большая часть глицерофосфата используется для синтеза липидов. Часть глицерофосфата окисляется до фосфодиоксиацетона, который изомеризуется в глицеральдегид-3-фосфат, который является промежуточным продуктом гликолиза и используется клеткой для получения энергии (рис. 16).
Жирные кислоты являются важным продуктом катаболических путей. Первым этапом распада жирных кислот является их активирование, этот процесс происходит на внешней мембране митохондрий и мембранах эндоплазматической сети и катализируется ферментом ацил-КоА-синтетазой. В результате происходит присоединение остатка жирной кислоты к Коферменту А.
В митохондриях происходит процесс окисления жирных кислот, который представляет собой постепенное уменьшение на два углеродных атома остова жирной кислоты. Этот процесс носит название b-окисления, его схема представлена на рис. 17.
Рис. 17. b-Окисление жирных кислот
Первой стадией b-окисления жирных кислот является окисление ацил-КоА путем отщепления двух атомов водорода от a- и b-углеродных атомов ацильного остатка в составе ацил-КоА. Далее происходит присоединение молекулы воды по двойной связи образовавшегося еноил-КоА. Затем происходит следующая реакция окисления, продуктом которой является кетоацил-КоА. На последующей стадии b-окисления происходит распад кетоацила-КоА и перенос ацильного остатка, укороченного на два углеродных атома по сравнению с первоначальным, на молекулу КоА. Продуктом этой реакции также является ацетил-КоА.
Читайте также: В куске 36м ткани
Образовавшийся ацил-КоА вновь подвергается b-окислению по той же схеме, соответственно имеет место циклический процесс, продолжающийся до тех пор пока от длинного остова жирной кислоты не останется двухуглеродного фрагмента. Таким образом, конечным продуктом b-окисления жирных кислот является ацетил-КоА, дальнейшая судьба которого зависит от состояния организма. В том случае, например, когда организму необходима энергия, он будет окисляться в цикле Кребса, как обсуждалось выше.
Образование кетоновых тел
Для протекания процессов окисления жирных кислот необходимо наличие свободного кофермента А. Одним из процессов, в котором происходит регенерирование свободного КоА из его ацильных производных, является образование ацетоуксусной кислоты. В этом процессе принимают участие три молекулы ацетил-КоА (рис. 18).

Рис. 18. Образование кетоновых тел
Сначала происходит конденсация двух молекул ацетил-КоА с образованием b-кетобутирил-КоА. Далее имеет место высвобождение КоА из b-кетобутирил-КоА с использованием еще одной молекулы ацетил-КоА, что приводит к образованию b-окси-b-метилглутарил-КоА. Последнее соединение подвергается расщеплению, в результате которого образуется ацетоуксусная кислота. Таким образом, в результате конденсации трех молекул ацетил-КоА образуется молекула ацетоуксусной кислоты и высвобождается две молекулы КоА. Ацетоуксусная кислота может восстанавливаться с образованием b-оксимасляной кислоты, либо декарбокисилироваться с образованием ацетона.
Ацетоуксусная и b-оксимасляная кислота синтезируются в печени и поступают с током крови к мышечной и другим тканям, где они могут утилизироваться в цикле Кребса. Ацетоуксусная, b-оксимасляная кислоты и ацетон получили название кетоновых тел. Усиленное их образование называется кетозом. Состояние организма, при котором происходит избыточное накопление кетоновых тел в крови, называется кетонемией, а выделение их с мочой – кетонурией. Среди многих причин патологического накопления кетоновых тел особенно важным является дефицит поступающих с пищей углеводов и преимущественное использование жиров в катаболических процессах, а также нарушение обмена углеводов и жиров при недостатке гормона инсулина. Избыточное содержание кетоновых тел в крови оказывает токсическое действие на организм.
Анаболизм липидов
Основными структурными блоками нейтральных жиров и фосфолипидов являются Глицерофосфат и ацильные производные Кофермента А (ацил-КоА). Глицерофосфат образуется из глицерина, возникающего при распаде глицеринсодержающих липидов, а высшие жирные кислоты синтезируются из малонил-КоА.
Синтез жирных кислот
Синтез жирных кислот локализован на мембранах гладкого эндоплазматического ретикулума. Исходным продуктом этого синтеза является малонил-КоА, который образуется при карбоксилировании ацетил-КоА (рис. 19). Эта реакция называется гетеротрофной фиксацией СО2.

Рис. 19. Синтез жирных кислот
Начальным этапом синтез жирных кислот является конденсация малонил-КоА с ацетил-КоА, в результате которой образуется кетобутирил-КоА. Кетобутирил-КоА восстанавливается до оксибутирил-КоА, который далее дегидратируется с образованием кротонил-КоА. Кротонил-КоА восстанавливается до бутирил-КоА. А далее происходит конденсация образовавшегося бутирил-КоА со следующим ацетил-КоА (рис. 20).
Таким образом, синтез жирных кислот носит циклический характер и представляет собой последовательность присоединений двухуглеродного остатка к растущей цепи с последующим восстановлением продукта конденсации. После того, как ацильный радикал достигает нужного размера, он отщепляется от Кофермента А с помощью специального фермента.
Биосинтез нейтральных жиров и фосфолипидов
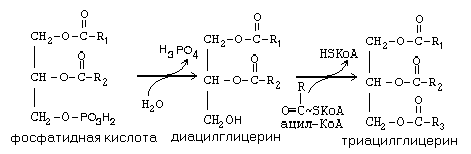
Синтез липидов происходит в гладкой эндоплазматической сети. Сначала происходит присоединение двух ацильных остатков к молекуле глицерофосфата, в результате этого процесса образуется фосфатидная кислота, которая является общим предшественником и нейтральных жиров, и фосфолипидов (рис. 20).
Рис. 20. Биосинтез нейтральных жиров и фосфолипидов
Далее, в ходе образования нейтральных жиров происходит отщепление фосфата от молекулы фосфатидной кислоты, образовавшийся при этом диацилглицерин еще раз ацилируется опять же с участием ацил-КоА и образуется нейтральный липид (триглицерид). Активный синтез нейтральных липидов происходит в печени, слизистой оболочке кишечника, жировой ткани. Фосфатидная кислота служит предшественником при синтезе фосфолипидов. При этом происходит присоединение соответствующего радикала к остатку фосфорной кислоты в составе фосфатидной кислоты. Этот процесс может включать несколько промежуточных стадий, которые зависят, в первую очередь, от природы присоединяемого радикала.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом