
Вопрос 49.Биосинтез таг (липогенез). Особенности биосинтеза таг в печени и жировой ткани. Гормональная регуляция. Образование лпонп в печени.
Печень — основной орган, где идет синтез жирных кислот из продуктов гликолиза. В гладком Эр гепатоцитов жирные кислоты активируются и сразу же используются для синтеза жиров, взаимодействуя с глицерол-3-фосфатом. Как и в жировой ткани, синтез жиров идет через образование фосфатидной кислоты. Синтезированные в печени жиры упаковываются в ЛПОНП и секретируются в кровь.В состав ЛПОНП, кроме жиров, входят холестерол, фосфолипиды и белок апоВ-100. Это очень «длинный» белок , содержащий 11536 аминокислот. Одна молекула апоВ-100 покрывает поверхность всего липопротеина. ЛПОНП из печени секретируется в кровь, где на них, как и на ХМ, действует ЛП-липаза. Жирные кислоты поступают в ткани, в частности в адипоциты, и используются для синтеза жиров. В процессе удаления жиров из ЛПОНП под действием ЛП-липазы ЛПОНП сначала превращаются в ЛППП , а затем в ЛПНП. В ЛПНП основными липидными компонентами служат холестерол и его эфиры, поэтому ЛПНП являются липопротеинами, доставляющими холестерол в периферические ткани. Глицерол, освободившийся из липопротеинов, кровью транспортируется в печень, где опять может использоваться для синтеза жиров. Скорость синтеза ЖК и жиров в печени существенно зависит от состава пищи. Если в пище содержится белее 10% жиров, то скорость синтеза жиров в печени резко снижается.
Гормональная регуляция синтеза жиров. Какой процесс будет преобладать в организме – синтез жиров или их распад зависит от поступления пищи и физической активности В абсорбтивном состоянии под действием инсулина происходит липогенез, в постабсорбтивном состоянии – липолиз, активируемый глюкагоном. Адреналин, секреция которого увеличивается при физической активности, также стимулирует липолиз. В абсорбтивный период при увеличении соотношения инсулин/глюкагон в печени активируется синтез жиров. В жировой ткани индуцируется синтез ЛП-липазы в адипоцитах и осуществляется ее экспонирование на поверхность эндотелия , следовательно в этот период увеличивается поступление жирных кислот в адипоциты. Одновременно инсулин активирует белки-переносчики глюкозы – ГЛЮТ-4. Поступление глюкозы в адипоциты и гликолиз также активируется. В результате образуются все необходимые компоненты для синтеза жиров: глицерол-3-фосфат и активные формы жирных кислот. В печени инсулин, действуя через различные механизмы, активирует ферменты путем дефосфорилирования и индуцирует их синтез. В результате увеличиваются активность и синтез ферментов, участвующих в превращении части глюкозы, поступающей с пищей в жиры. Это – регуляторные ферменты гликолиза, пируватдегидрогеназный комплекс и ферменты, участвующие в синтезе ЖК из ацетил-КоА. Результат действия инсулина на обмен углеводов и жиров в печени – увеличение синтеза жиров и секреция их в кровь в составе ЛПОНП. ЛПОНП доставляют жиры в капилляры жировой ткани, где действие ЛП-липазы обеспечивает быстрое поступление жирных кислот в адипоциты, где они депонируются в составе ТАГ.

Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Синтез жиров (ТАГ)
Обмен жиров или ТАГ включает в себя несколько стадий: 1). Синтез жиров (из глюкозы, эндогенные жиры), 2). Депонирование жиров, 3). Мобилизация.
В организме жиры могут синтезироваться из глицерина и из глюкозы. Основные 2 субстрата для синтеза жиров:
2) ацилКоА (активированная ЖК).
Синтез ТАГ происходит через образование фосфатидной кислоты.
α-ГФ в организме человека может образовываться двумя путями: в органах, в которых активен фермент глицеролкиназа, ГФ может образоваться из глицерина, в органах, где активность фермента низкая, ГФ образуется из продуктов гликолиза (т.е. из глюкозы).
Если в реакцию вступает восстановленная форма НАД (НАДН+Н ), то это реакция
восстановления и фермент называется по продукту + «ДГ».
Биосинтез ТАГ наиболее интенсивно протекает в печени и жировой ткани. В жировой
ткани синтез ТАГ протекает из УВ, т.е. часть глюкозы, поступившей с пищей может
превращаться в жиры (когда углеводов поступает больше, чем необходимо для
возобновления запаса гликогена в печени и мышцах).
Жиры, синтезированные в печени (двумя путями) упаковываются в частицы ЛОИП,
поступают в кровь —> ЛП-липазе, которая гидролизует ТАГ или жиры из этих частиц на
ЖК и глицерин. ЖК поступают в жировую ткань, где депонируются в виде жиров, либо
используются как источник энергии органами и тканями (р-окисление), а глицерин
поступает в печень, где может использоваться для синтеза ТАГ или фосфолипидов.
В жировой ткани депонируются жиры, которые образованы из глюкозы, глюкоза дает
оба или 2 субстрата для синтеза жира.
После приема пищи (абсорбционный период) f концентрация глюкозы в крови, |
концентрация инсулина, инсулин активирует:
1. транспорт глюкозы в адипоциты,
—► активирует синтез жира в жировой ткани и его депонирование —> существует 2 источника жиров для депонирования в жировой ткани:
1. экзогенные (ТАГ из хиломикронов и ЛОНП кишечника, переносящие пищевые
жиры)
2. эндогенные жиры (из ЛОНП печени и образующиеся ТАГ в самих жировых
клетках).
Мобилизация жиров — это гидролиз жиров, находящихся в адипоцитах до ЖК и глицерина, под действием гормонзависимой ТАГ-липазы, которая находится в клетках и активируется в зависимости от потребностей организма в источниках энергии (в постабсорбтивном периоде, т.е. в промежутках между приемами пищи, при голодании, стрессе, длительной физической работе, т.е. активируется адреналином, глюкагоном и соматотропным гормоном (СТГ).
Читайте также: Как называется ткань для накрытия грядок
При длительном голодании концентрация глюкагона увел., это приводит к снижению синтеза ЖК, увеличению β-окисления, увеличеню мобилизации жиров из депо, увеличен синтез кетоновых тел, увеличен глюконеогенез.
Отличие действия инсулина в жировой ткани и печени:
↑ концентрации инсулина в крови приводит к ↑ активности ПФП, ↑ синтеза ЖК, ↑ гликолиза (глюкокиназа, фосфофруктокиназа (ФФК), пируваткиназа — ферменты гликолиза; глюкозо-6-ДГ — фермент ПФП; ацетилКоАкарбоксилаза — фермент синтеза ЖК).
В жировой ткани активируется ЛП-липаза и депонирование жиров, активируется поступление глюкозы в адипоциты и образование из нее жиров, которые тоже депонируются.
В организме человека существует 2 формы депонированного энергетического материала:
1. гликоген; 2. ТАГ или нейтральные жиры.
Отличаются по запасам и очередности мобилизации. Гликогена в печени от 120-150г, может быть до 200, жиров в норме
Гликогена хватает (в качестве источника энергии) на 1 сутки голодания, а жиров — на 5-7 недель.
При голодании и физической нагрузке в первую очередь используются запасы гликогена, затем постепенно нарастает скорость мобилизации жиров. Кратковременные физические
нагрузки обеспечиваются энергией, за счет распада гликогена, а при длительных физических нагрузках используются жиры.
При нормальном питании количество жиров в жировой ткани постоянно, но жиры постоянно обновляются. При длительном голодании и физических нагрузках скорость мобилизации жиров больше, чем скорость депонирования à количество депонированных жиров уменьш. (похудение). Если скорость мобилизации ниже, чем скорость депонирования — ожирение.
Причины: несоответствие между количеством потребляемой пищи и энергозатратами организма, и поскольку мобилизация и депонирование жиров регулируются гормонами —» ожирение является характерным признаком эндокринных заболеваний.
Обмен холестерина. Биохимические основы возникновения атеросклероза. Основные функции холестерина в организме:
1. основная: большая часть Хс используется для построения клеточных мембран;
2. Хс служит предшественником желчных кислот;
3. служит предшественником стероидных гормонов и витамина D3 (половые
гормоны и гормоны коры надпочечников).
В организме на долю Хс приходится основная масса всех стероидов
140г. Синтезируется Хс в основном в печени (-80%), в тонком кишечнике (-10%), в коже (-5%), скорость синтеза Хс в организме зависит от количества экзогенного Хс, если с пищей поступает более 1г Хс (2-3г) синтез собственного эндогенного Хс ингибируется, если Хс поступает мало (вегетарианцы) скорость синтеза эндогенного Хс |. Нарушение в регуляции синтеза Хс (а также образование его транспортных форм —> гиперхолестеринемия —» атеросклероз —> ИБС —► инфаркт миокарда). Норма поступления Хс >1г (яйца, масло (сливочное), печень, мозг).
Синтез ТАГ — это запасание энергии
Липогенез
Синтез триацилглицеролов (липогенез) заключается в дефосфорилировании фосфатидной кислоты, полученной из глицерол-3-фосфата, и присоединении ацильной группы.
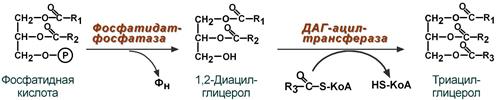
Реакции синтеза ТАГ из фосфатидной кислоты
Если синтез ТАГ происходил в печени, то они эвакуируются из нее в ткани, имеющие на эндотелии своих капилляров липопротеинлипазу (транспорт ТАГ в крови). Транспортной формой служат ЛПОНП. Строго говоря, клеткам организма нужны только жирные кислоты, все остальные компоненты ЛПОНП не являются необходимыми.
Синтез ТАГ увеличивается при соблюдении хотя бы одного из следующих условий, которые обеспечивают появление избытка ацетил-SКоА :
- наличие источника «дешевой» энергии . Например,
1) диета богатая простыми углеводами (глюкоза, сахароза). При этом концентрация глюкозы в печени и адипоцитах после еды резко повышается, она окисляется до ацетил-SКоА и под влиянием инсулина в этих органах активно происходит синтез жиров.
2) наличие этанола, высокоэнергетичного соединения, который окисляется до ацетил-SКоА. «Алкогольный» ацетил используется в печени для синтеза жира при условии нормального питания. Примером может служить «пивное ожирение». - повышение концентрации жирных кислот в крови . Например, при усиленном липолизе в жировых клетках под воздействием каких-либо веществ (фармпрепараты, кофеин и т.п.), при эмоциональном стрессе и отсутствии (!) мышечной активности увеличивается поток жирных кислот в гепатоциты. Здесь в результате происходит интенсивный синтез ТАГ.
- высокие концентрации инсулина и низкие концентрации глюкагона – после приема высокоуглеводной и жирной пищи.
Синтез фосфолипидов и триацилглицеролов тесно связаны
Начальные реакции синтеза триацилглицеролов и фосфолипидов совпадают и происходят при наличии глицерола и жирных кислот .
В реакциях биосинтеза можно выделить следующие события:
1. Образование глицерол-3-фосфата через диоксиацетонфосфат из глюкозы или при фосфорилировании свободного глицерола.
2.. Биосинтез фосфатидной кислоты – требует наличия глицерол-3-фосфата и жирных кислот. При связывании глицерол-3-фосфата с жирными кислотами синтезируется фосфатидная кислота.
Далее фосфатидная кислота может превращаться двумя путями – в ЦДФ-ДАГ или дефосфорилироваться до 1,2-ДАГ.
3. Синтез триацилглицерола (липогенез) – идет из 1,2-ДАГ после дефосфорилирования фосфатидной кислоты. Образованный 1,2-ДАГ ацилируется до ТАГ.
4. Синтез фосфолипидов . Сейчас рассматриваются два пути синтеза фосфолипидов.
- По одному пути 1,2-ДАГ не превращается в ТАГ, а связывается с этаноламином с образованием фосфатидилэтаноламина, либо с холином – образуется фосфатидилхолин.
- По другому пути, ЦДФ-ДАГ связывается либо с инозитолом, либо с серином с образованием соответственно фосфатидилинозитола или фосфатидилсерина. При декарбоксилировании фосфатидилсерина далее образуется фосфатидилэтаноламин, который может превратиться, в свою очередь, в фосфатидилхолин.
Читайте также: Ткань с принтом для кухни
Синтезированный любым способом фосфатидилэтаноламин также способен взаимодействовать с серином и обратно образовывать фосфатидилсерин.
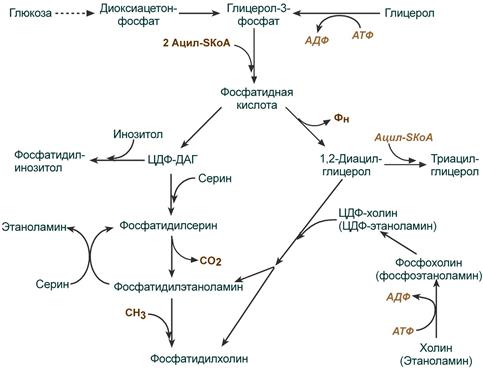
Общая схема реакций синтеза триацилглицеролов и фосфолипидов
ЦДФ-ДАГ, являясь активной формой фосфатидной кислоты, способен превращаться не только в фосфатидилинозитол, фосфатидилсерин, но и в другие фосфолипиды, например в кардиолипин .
5_Lektsia_Metabolizm_TAG_i_FL
Основным местом накопления ТАГ является жировая ткань (адипоциты).
Жировые клетки выполняют три функции:
3) ВЖК, находящиеся в составе ТАГ, используются для получения энергии.
Катаболизм ТАГ осуществляется под действием ТАГ-липазы. Конечным продуктом ферментативного гидролиза ТАГ являются: жирные кислоты и глицерол.
Катаболизм ТАГ в адипоцитах и судьба продуктов гидролиза
Освободившиеся ЖК поступают в плазму, связываются с альбуминами крови и доставляются в периферические ткани, где они либо окисляются, либо идут вновь для синтеза липидов.
Глицерол, образовавшийся при липолизе, поступает из жировой ткани в кровь и активно утилизируется тканями, в которых активна глицеролкиназа (печень,
почки, кишечник, лактирующая молочная железа). Там глицерол фосфорилируется под действием глицеролкиназы, образуется α-глицерофосфата
(глицерол-3-фосфата), который окисляется в фосфодигидроксиацетон (ДГАФ)
под действием НАД-зависимой глицерол-3-фосфатдегидрогеназы (реакция обратимая).
Далее ДГАФ изомеризуется под действием триозофосфатизомеразы в 3-
фосфоглицероловый альдегид (3-ФГА). 3-ФГА – это промежуточный продукт и гликолиза, и глюконеогенеза. Следовательно, из него может образовываться и ПВК, и глюкоза.
Возможен также и обратный процесс — образование из глюкозы ДГАФ, его дальнейшее восстановление в α-глицерофосфат (Е – глицерол-3-фосфат ДГ,
реакция обратима), а затем гидролиз α-ГФ под действием глицеролфосфатазы с образованием глицерола.
В животных тканях для биосинтеза ТАГ необходимы ацил-KoA и α-ГФ.
α-ГФ может быть образован из:
1) глицерола при участии глицеролкиназы и АТФ, этот глицеролкиназный путь синтеза α-ГФ активен в тех тканях, где высока активность глицеролкиназы
(печень, почки, кишечник, молочная железа); 2) ДГАФ – метаболита гликолиза, который затем в ходе обратимых реакций
превращается в α-ГФ. Такой путь характерен для мышц и жировой ткани, так как в них отсутствует глицеролкиназа или активность фермента очень низка. АцилKoA образуется из ацетил-KoA.
В биосинтезе ТАГ выделяют три этапа
1. Этерификация (ацилирование) свободных НО-групп α-ГФ двумя молекулами ацил-KoA с образованием фосфатидной кислоты (фермент – ацилтрансфераза или трансацилаза)
2. Гидролиз фосфатидной кислоты с образованием 1,2-ДАГ (фермент –
3. Ацилирование 1,2-ДАГ и образование ТАГ (фермент – ацилтрансфераза).
Для энтероцитов более характерен ресинтез ТАГ из β-МАГ (фермент – β-МАГ-
Таким образом, субстратами для синтеза ТАГ являются:
1. α-ГФ (из глюкозы или глицерола);
3. В клетках кишечника – β-МАГ.
Катаболизм ФЛ в клетке (Не путать с полостным перевариванием в кишечнике!). При полном распаде ФЛ под действием клеточных фосфолипаз А 1 ,
А 2 , С, D, локализованных в мембранах или лизосомах, образуются: 1) ВЖК; 2)
глицерол; 3) Н 3 РО 4 ; 4) аминоспирт.
Синтез мембранных липидов (ФЛ)
Основные липиды клеточных мембран – это фосфатидилэтаноламин (ФЭА) и
фосфатидилхолин (ФХ). Они образуются, так же как и ТАГ, из общего промежуточного соединения – 1,2-ДАГ. Для того, чтобы 1,2-ДАГ превратились в ФЛ, им необходимо приобрести группы, образующие «полярные головки»: Н 3 РО 4
и аминоспирт. Процесс образования ФЛ связан с образованием активных форм аминоспиртов. Для этого используются два макроэрга – АТФ и ЦТФ.
ФХ в организме человека может быть синтезирован двумя путями: 1)
основной путь синтезом «de novo» из фосфатидилэтаноламина, и 2) утилизацией экзогенного холина из пищи.
В первом случае экзогенный холин не требуется. Он образуется в процессе трёхкратного метилирования ФЭА. Донор метильных групп – активная форма метионина – S-аденозилметионин (SАМ), переходящий в S-аденозилгомоцис-
теин. Этот путь может быть блокировании или ограничен при недостатке метионина.
Фосфатидилсерин (ФС) синтезируется путём прямого взаимодействия ФЭА и серина. ФС может декарбоксилироваться с образованием ФЭА.
Путь синтеза ФХ de novo не может покрыть все потребности в нем организма, поэтому необходим экзогенный источник холина и других азотсодержащих компонентов фосфолипидов. В этом случае в синтезе ФЛ используются
активные формы этаноламина – фосфоэтаноламин, холина – фосфохолин.
Сначала этаноламин фосфорилируется в реакции с АТФ и образуется фосфоэтаноламин (фермент – этаноламинкиназа). Холин аналогичным образом
переходит в фосфохолин (фермент – холинкиназа). Затем в реакции с ЦТФ образуется цитидилдифосфоэтаноламин (ЦДФ-ЭА) и, соответственно,
цитидилдифосфохолин (ЦДФ-ФХ) – это активные формы аминоспиртов.
Катализирует их синтез регуляторный фермент – цитидилтрансфераза.
Образовавшиеся активные формы переносят остатки фосфоэтаноламина и фосфохолина на ДАГ (т.е. происходит перенос полярной «головы» —
фосфоэтаноламина (ФЭА) и фосфохолина (ФХ) на 1,2-ДАГ) с образованием ФЭА
Читайте также: Обои из кусочков ткани своими руками
и ФХ. Катализируют перенос регуляторные ферменты –
фосфоэтаноламинтрансфераза и фосфохолинтрансфераза.
Рассмотренный путь синтеза ФЛ называется «спасательным или резервным», так как в нём используется готовый холин. Таким образом, холин сохраняется от разрушения путём реутилизации.
Регуляторными ферментами являются:
1. Цитидилтрансфераза – фермент локализован в цитозоле, активность увеличивается при высоких концентрациях холина и этаноламина и, наоборот,
снижается при низких концентрациях этих субстратов.
2. Фосфоэтаноламин- и фосфохолинтрансфераза . Эти ферменты локализованы в цитозоле. Их активность увеличивается в присутствии ДАГ и снижается в их отсутствии. Это саморегулирующаяся система.
Симпатическая нервная система активирует синтез ФХ, поставляя в систему синтеза фосфолипидов ацетилхолин.
Регуляция синтеза и распада ТАГ в печени и жировой ткани
В период приёма пищи в печени и жировой ткани под действием инсулина происходит липогенез (синтез ТАГ). Для осуществления липогенеза необходим ацетил-KoA. Инсулин стимулирует утилизацию глюкозы печенью за счет индукции глюкокинды. В печень поступает много глюкозы, в результате чего активируется гликолиз и ЦТК. В результате накапливающийся цитрат переходит в цитозоль, расщепляется на исходные компоненты: ацетил-КоА и
ОАА, и активирует регуляторный фермент ацетил-КоА-карбоксилазу,
активирующую синтез малонил-КоА. Малонил-КоА блокирует активность регуляторного фермента β-окисления жирных кислот (ЖК) –
карнитинацилтрансферазу I, а сам активно включается в биосинтез жирных кислот (ЖК).
В результате действия инсулина увеличивается синтез ТАГ и секреция их в кровь в составе ЛПОНП. ЛПОНП доставляют жиры (ТАГ) из печени в капилляры жировой ткани. В жировой ткани инсулин индуцирует синтез липопротеид-липазы. ЛП-липаза переносится на поверхность эндотелия капилляров жировой ткани, где она осуществляет гидролиз ТАГ, находящихся в составе ЛПОНП. Следовательно, увеличивается поступление жирных кислот в адипоциты (ТАГ распадается до 1,2-ДАГ + ВЖК (высшие жирные кислоты), т.к.
ЛП-липаза действует в 1 или 3 положениях).
Одновременно инсулин активирует белок-переносчик глюкозы глют-4 в
адипоцитах. Поступление глюкозы в адипоциты усиливается и активируется гликолиз. В процессе гликолиза образуются все необходимые компоненты для синтеза ТАГ: α-ГФ, активные формы жирных кислот (ацил-КоА).
1. Между приёмами пищи, при голодании и физической нагрузке активируется липолиз (распад жиров), который стимулируется глюкагоном и адреналином и, в меньшей степени, кортизолом и соматотропным гормоном.
2. Гормоны адреналин и глюкагон, действуя на адипоциты через аденилатциклазную сисему, активируют протеинкиназу А. Протеинкиназа А фосфорилирует ТАГ-липазу (активна в фосфорилированной форме). ТАГ-липаза расщепляет ТАГ на глицерол и ВЖК, которые выделяются в кровь и доставляются альбуминами в мышцы, сердце, печень и почки. Печень перерабатывает часть ЖК в кетоновые тела, используемые мозгом, сердцем,
мышцами, корковым слоем почек как источники энергии.
3. Во время приёма пищи инсулин активирует специфическую фосфатазу,
которая дефосфорилирует ТАГ-липазу и распад жиров останавливается, так как
ТАГ-липаза в дефосфорилированном виде неактивна.
Регуляция обмена ТАГ в печени и жировой ткани
Клинические аспекты обмена мембранных липидов
Полярные липиды не запасаются в жировых клетках подобно ТАГ, а
встраиваются в различные типы клеточных мембран.
1. Дипальмитоилфосфатидилхолин (лецитин) – (ФХ, содержащий два остатка С 16:0 ) – основной компонент сурфактанта лёгких.
Сурфактант – внеклеточный липидный слой с небольшим количеством гидрофобных белков, выстилающий поверхность легочных альвеол и предотвращающий слипание стенок альвеол во время выдоха и перерастяжение их во время вдоха. На долю дипальмитоилфосфатидилхолина приходится 80% от всех фосфолипиды,
входящих в состав сурфактанта. Кроме того, компонентами сурфактанта являются гидрофобные белки, доля которых составляет 10-20% от общей массы сурфактанта.
Синтез дипальмитоилфосфатидилхолина происходит в пневмоцитах II типа в период с 32 по 36 недели беременности. Показателем нормального формирования сурфактанта служит величина соотношения фосфатидилхолин/сфингомиелин более 4. Снижение этого коэффициента до значений менее 2 указывает на высокий риск развития респираторного дистресс-синдрома, который наблюдается у недоношенных детей. В случае необходимости беременных лечат кортикостероидными препаратами,
которые стимулируют синтез сурфактанта.
2. Недостаток полярных липидов может быть причиной другого патологического состояния – рассеянного склероза . При этом заболевании наблюдается снижение содержания фосфолипидов и сфинголипидов в белом веществе мозга. В большом количестве присутствуют эфиры ХСа,
отсутствующие в норме, а спинномозговая жидкость обогащается фосфолипидами.
Сурфактант уменьшает поверхностное натяжение жидкости, выстилающей поверхность альвеол и предотвращающей слипание стенок альвеол во время выдоха. Меньшее давление воздуха необходимо, чтобы
наполнить альвеолы воздухом.
Альвеолы в момент выдоха при недостатке сурфактанта .
Отсутствие или недостаток сурфактанта (у недоношенных детей) приводит к тому, что стенки альвеол во время выдоха спадаются и требуют давление воздуха в 10 раз больше, чтобы наполнить альвеолы .
3. Полярные липиды в мембранах постоянно обновляются. Их
расщепление происходит в лизосомах под действием соответствующих
гидролаз. Известны генетические заболевания, связанные с дефектами
ферментов, участвующих в деградации липидов, под общим названием
лизосомные болезни . Продукты неполного расщепления липидов
накапливаются в лизосомах, которые сильно разбухают, что приводит к
Болезнь Тея-Сакса связана с дефектом гексозаминидазы , которая
гидролизует связь между галактозамином и галактозой в полярной головке
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом