
Мышечное сокращение является жизненно важной функцией организма, связанной с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами. Все виды произвольных движений – ходьба, мимика, движения глазных яблок, глотание, дыхание и т. п. осуществляются за счет скелетных мышц. Непроизвольные движения (кроме сокращения сердца) – перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря – обусловлены сокращением гладких мышц. Работа сердца обеспечивается сокращением сердечной мускулатуры.
Структурная организация скелетной мышцы
Мышечное волокно и миофибрилла (рис. 1). Скелетная мышца состоит из множества мышечных волокон, имеющих точки прикрепления к костям и расположенных параллельно друг другу. Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл , которые построены из повторяющихся в продольном направлении блоков (саркомеров). Саркомер является функциональной единицей сократительного аппарата скелетной мышцы. Миофибриллы в мышечном волокне лежат таким образом, что расположение саркомеров в них совпадает. Это создает картину поперечной исчерченности.
Рис. 1. Иерархические уровни организации скелетной мышцы (по Bloom , 1968).
Саркомер и филламенты. Саркомеры в миофибрилле отделены друг от друга Z -пластинками, которые содержат белок бета-актинин. В обоих направлениях от Z -пластинки отходят тонкие актиновые филламенты. В промежутках между ними располагаются более толстые миозиновые филламенты .
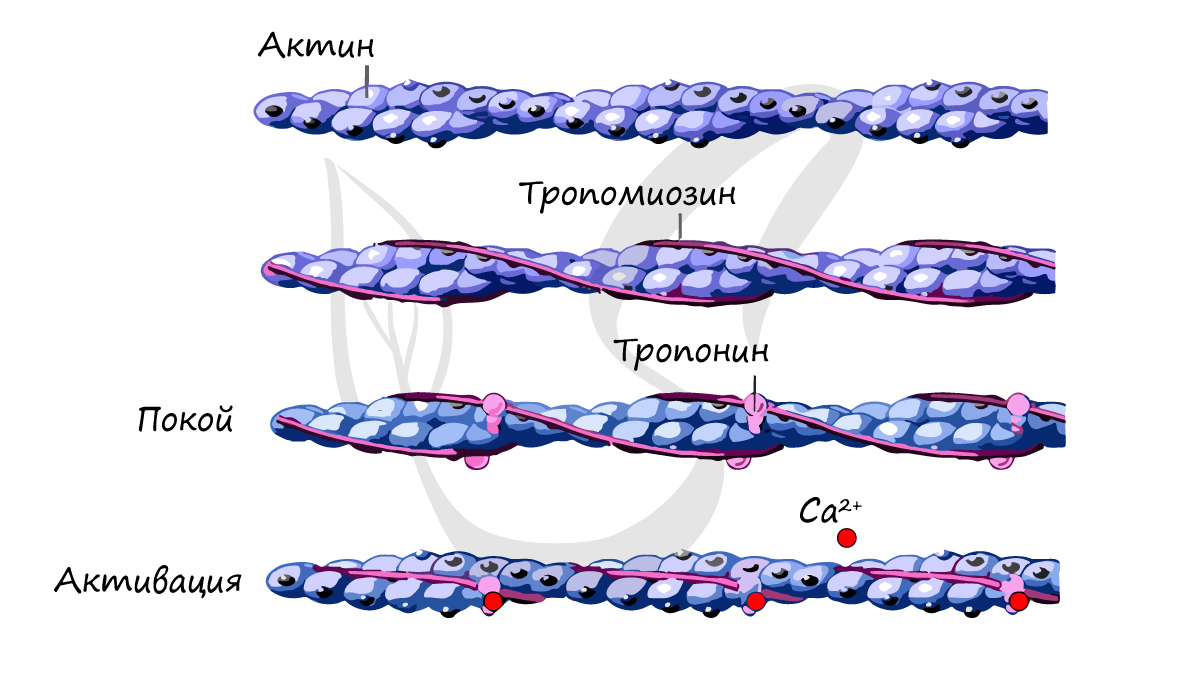
Актиновый филламент внешне напоминает две нитки бус, закрученные в двойную спираль, где каждая бусина – молекула белка актина . В углублениях актиновых спиралей на равном расстоянии друг от друга лежат молекулы белка тропонина , соединенные с нитевидными молекулами белка тропомиозина.
Миозиновые филламенты образованы повторяющимися молекулами белка миозина . Каждая молекула миозина имеет головку и хвост . Головка миозина может связываться с молекулой актина, образуя так называемый поперечный мостик .
Клеточная мембрана мышечного волокна образует инвагинации (поперечные трубочки), которые выполняют функцию проведения возбуждения к мембране саркоплазматического ретикулума. Саркоплазматичекий ретикулум (продольные трубочки) представляет собой внутриклеточную сеть замкнутых трубочек и выполняет функцию депонирования ионов Са++ .
Двигательная единица. Функциональной единицей скелетной мышцы является двигательная единица (ДЕ). ДЕ – совокупность мышечных волокон, которые иннервируются отростками одного мотонейрона. Возбуждение и сокращение волокон, входящих в состав одной ДЕ, происходит одновременно (при возбуждении соответствующего мотонейрона). Отдельные ДЕ могут возбуждаться и сокращаться независимо друг от друга.
Молекулярные механизмы сокращения скелетной мышцы
Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга. Механизм скольжения нитей включает несколько последовательных событий.
Головки миозина присоединяются к центрам связывания актинового филламента (рис. 2, А).
Взаимодействие миозина с актином приводит к конформационным перестройкам молекулы миозина. Головки приобретают АТФазную активность и поворачиваются на 120 ° . За счет поворота головок нити актина и миозина передвигаются на «один шаг» друг относительно друга (рис. 2, Б).
Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина молекулы АТФ и ее гидролиза в присутствии Са++ (рис. 2, В).
Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются и миофибрилла укорачивается (рис. 2, Г).
Рис. 2. Механизм мышечного сокращения.
Объяснение – в тексте.
1 – актиновый филламент,
2 – центр связывания,
3 – миозиновый филламент,
4 – головка миозина,
5 – Z -диск саркомера.
Сопряжение возбуждения и сокращения в скелетной мышце
В состоянии покоя скольжения нитей в миофибрилле не происходит, так как центры связывания на поверхности актина закрыты молекулами белка тропомиозина (рис. 3, А, Б). Возбуждение (деполяризация) миофибриллы и собственно мышечное сокращение связаны с процессом элетромеханического сопряжения, который включает ряд последовательных событий.
Рис. 3. Механизм сопряжения возбуждения и сокращения.
Объяснение – в тексте.
1 – поперечная трубочка саркоплазматичекой мембраны,
2 –саркоплазматичекий ретикулум,
3 – ион Са++ ,
4 – молекула тропонина,
5 – молекула тропомиозина.
В результате срабатывания нейромышечного синапса на постсинаптической мембране возникает ВПСП, который генерирует развитие потенциала действия в области, окружающей постсинаптическую мембрану.
Возбуждение (потенциал действия) распространяется по мембране миофибриллы и за счет системы поперечных трубочек достигает саркоплазматического ретикулума. Деполяризации мембраны саркоплазматического ретикулума приводит к открытию в ней Са++ -каналов, через которые в саркоплазму выходят ионы Са++ (рис. 3, В).
Ионы Са++ связываются с белком тропонином. Тропонин изменяет свою конформацию и смещает молекулы белка тропомиозина, которые закрывали центры связывания актина (рис. 3, Г).
К открывшимся центрам связывания присоединяются головки миозина, и начинается процесс сокращения (рис. 3, Д).
Для развития указанных процессов требуется некоторый период времени (10–20 мс). Время от момента возбуждения мышечного волокна (мышцы) до начала ее сокращения называют латентным периодом сокращения .
Расслабление скелетной мышцы
Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере удаления Са++ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы.
Контрактурой называют стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя. Кратковременная контрактура может развиваться после тетанического сокращения в результате накопления в саркоплазме большого количества Са++ ; длительная (иногда необратимая) контрактура может возникать в результате отравления ядами, нарушений метаболизма.
Фазы и режимы сокращения скелетной мышцы
Фазы мышечного сокращения
При раздражении скелетной мышцы одиночным импульсом электрического тока сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы (рис. 4, А):
латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
фаза укорочения (около 50 мс);
фаза расслабления (около 50 мс).
Рис. 4. Характеристика одиночного мышечного сокращения. Происхождение зубчатого и гладкого тетануса.
Б – фазы и периоды иышечного сокращения,
Б – режимы мышечного сокращения, возникающие при разной частоте стимуляции мышцы.
Изменение длины мышцы показано синим цветом, потенциал действия в мышце — красным, возбудиумость мышцы — фиолетовым.
Режимы мышечного сокращения
В естественных условиях в организме одиночного мышечного сокращения не наблюдается, так как по двигательным нервам, иннервирующим мышцу, идут серии потенциалов действия. В зависимости от частоты приходящих к мышце нервных импульсов мышца может сокращаться в одном из трех режимов (рис. 4, Б).
Одиночные мышечные сокращения возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений.
При более высокой частоте импульсов очередной импульс может совпасть с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться, возникнет зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы.
При дальнейшем увеличении частоты импульсов каждый следующий импульс будет действовать на мышцу во время фазы укорочения, в результате чего возникнет гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления.
Оптимум и пессимум частоты
Амплитуда тетанического сокращения зависит от частоты импульсов, раздражающих мышцу. Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости (рис. 4, A) и соответственно вызывает тетанус наибольшей амплитуды. Пессимумом частоты называют более высокую частоту раздражения, при которой каждый последующий импульс тока попадает в фазу рефрактерности (рис. 4, A), в результате чего амплитуда тетануса значительно уменьшается.
Работа скелетной мышцы
Сила сокращения скелетной мышцы определяется 2 факторами:
числом ДЕ, участвующих в сокращении;
частотой сокращения мышечных волокон.
Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения.
Виды работы скелетной мышцы:
динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве;
статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении;
динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержать части тела.
Во время выполнения работы мышца может сокращаться:
изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте;
изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы;
ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе.
Правило средних нагрузок – мышца может совершить максимальную работу при средних нагрузках.
Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма. Утомляемость мышцы при ритмической работе меньше, чем утомляемость синапсов. Поэтому при совершении организмом мышечной работы утомление первоначально развивается на уровне синапсов ЦНС и нейро-мышечных синапсов.
Структурная организация и сокращение гладких мышц
Структурная организация . Гладкая мышца состоит из одиночных клеток веретенообразной формы (миоцитов), которые располагаются в мышце более или менее хаотично. Сократительные филламенты расположены нерегулярно, вследствие чего отсутствует поперечная исчерченность мышцы.
Механизм сокращения аналогичен таковому в скелетной мышце, но скорость скольжения филламентов и скорость гидролиза АТФ в 100–1000 раз ниже, чем в скелетной мускулатуре.
Механизм сопряжения возбуждения и сокращения . При возбуждении клетки Cа++ поступает в цитоплазму миоцита не только из саркоплазматичекого ретикулума, но и из межклеточного пространства. Ионы Cа++ при участии белка кальмодулина активируют фермент (киназу миозина), который переносит фосфатную группу с АТФ на миозин. Головки фосфорилированного миозина приобретают способность присоединяться к актиновым филламентам.
Сокращение и расслабление гладких мышц . Скорость удаления ионов Са++ из саркоплазмы значительно меньше, чем в скелетной мышце, вследствие чего расслабление происходит очень медленно. Гладкие мышцы совершают длительные тонические сокращения и медленные ритмические движения. Вследствие невысокой интенсивности гидролиза АТФ гладкие мышцы оптимально приспособлены для длительного сокращения, не приводящего к утомлению и большим энергозатратам.
Физиологические свойства мышц
Общими физиологическими свойствами скелетных и гладких мышц являются возбудимость и сократимость . Сравнительная характеристика скелетных и гладких мышц приведена в табл. 6.1. Физиологические свойства и особенности сердечной мускулатуры рассматриваются в разделе «Физиологические механизмы гомеостаза».
Таблица 7.1.Сравнительная характеристика скелетных и гладких мышц
Мышечные ткани
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.