
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
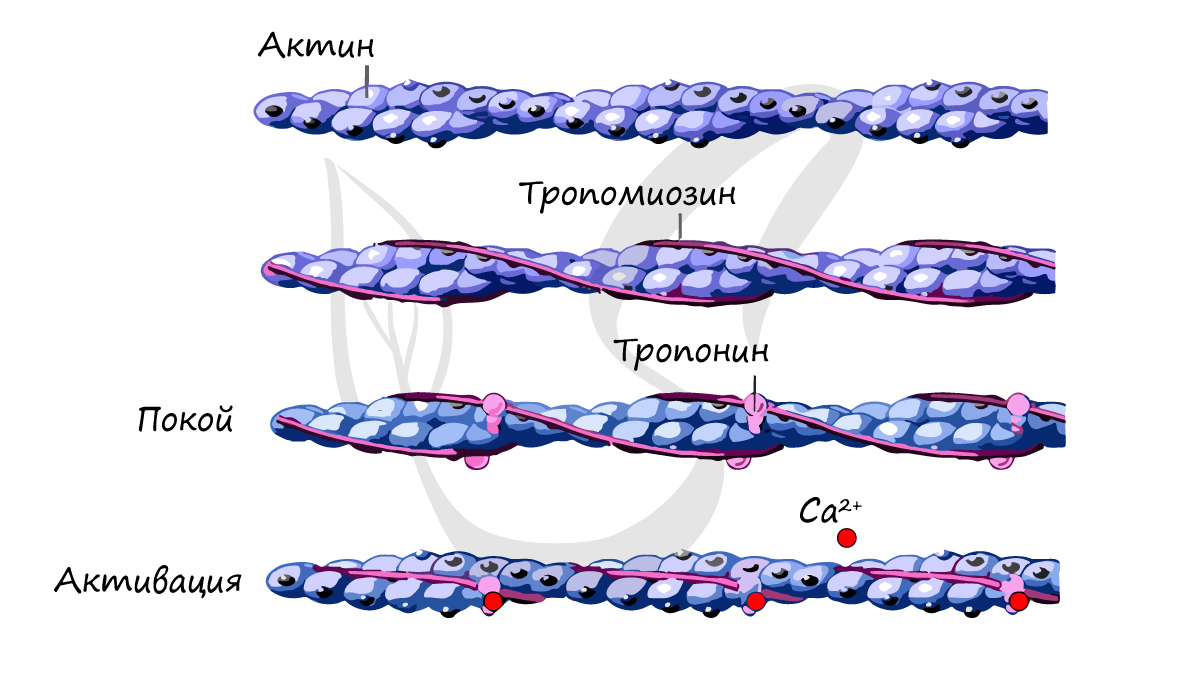
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.
Читайте также: Какая ткань нужна для мужских трусов

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Типы мышечных волокон
Типы волокон скелетных мышц [ править | править код ]

Мышечное волокно является структурной единицей мышечной ткани, которое состоит из:
- миофибрилл (сократительных элементов)
- митохондрий (энергопродукция)
- ядер (регуляция)
- сарколемы (соединительно-тканной оболочки)
- саркоплазматический или эндоплазматический ретикулум (депо кальция, необходимого для возбуждения миофибриллы)
- капилляры (поставка питательных веществ и кислорода)
У людей все волокна скелетных мышц имеют разные механические и метаболические свойства. Различные типы мышечных волокон определяют по максимальной скорости их сокращения (быстрой и медленной) и главного метаболического пути, который они используют для образования АТФ (окислительный и гликолитический). Мышечные волокна в целом делятся на:
- I тип: медленные окислительные (МО) — медленные, тонкие, слабые, неутомляемые мышечные волокна. Низкий порог активации мотонейрона. Волокна I типа хорошо кровоснабжаются и имеют большее количество миоглобина, что придает им характерный красный цвет (красные волокна). Они также отличаются наличием многочисленных крупных митохондрий, содержащих ферменты окислительного фосфорилирования. Хотя в медленных волокнах больше миозина, чем в быстрых мышечных волокнах, они содержат меньше фермента АТФазы и медленнее сокращаются. Иннервация обеспечивается малыми а-мотонейронами спинного мозга. Благодаря низкой скорости сокращения они больше приспособлены к длительным нагрузкам, что, например, очень важно для поддержания позы.
- II тип: быстрые гликолитические волокна — толще, чем мышечные волокна I типа, отличаются быстрыми сокращениями, развивают большую силу и быстрее утомляются. Эти волокна хуже кровоснабжаются и имеют меньше митохондрий, липидов и миоглобина. В литературе они описываются как белые волокна. В отличие от медленных волокон, быстрые волокна содержат в основном ферменты анаэробного окисления и больше миофибрилл. Эти миофибриллы отличаются меньшим содержанием миозина, который, однако, сокращается быстрее и лучше метаболизирует аденозинтрифосфат (АТФ). Кроме того, в этих волокнах лучше выражен саркоплазматический ретикулум. Благодаря высокой скорости сокращения и быстрой утомляемости эти волокна способны на кратковременную работу. Иннервация осуществляется большими а-мотонейронами спинного мозга. Эти волокна делятся на:
- IIа тип: быстрые окислительно-гликолитические (БОГ) или просто быстрые окислительные — промежуточные волокна, средней толщины. Более выносливы, чем волокна IIb типа, но утомляются быстрее, чем волокна I типа. Способны к выраженному сокращению, при этом развивают среднюю силу. Источниками энергии являются как окислительные, так анаэробные механизмы (быстрые окислительные волокна).
- IIb тип: быстрые гликолитические волокна (БГ) — крупные, быстрые, сильные, быстроутомляемые мышечные волокна, с высоким порогом активации мотонейрона. Активируются при кратковременных нагрузках и развивают большую силу. Получают энергию через процессы анаэробного окисления, источником энергии является гликоген. В этих волокнах обнаруживают большое количество гликогена и мало митохондрий.
Поскольку скорость сокращения самых быстрых мышечных волокон несколько выше, чем скорость сокращений волокон IIb типа, самые быстрые волокна называются в литературе волокнами IIх типа (Friedman, 2007).
Иногда выделяют волокна IIс типа — эти волокна не похожи на волокна ни I, ни II типа. Они проявляют как окислительную, так и гликолитическую активность и представлены лишь в небольшом количестве (около 1 %). В зависимости от типа тренировок они могут переходить в волокна I или II типа (Seidenspinner, 2005).
Мышечные волокна возбуждаемые одним мотонейроном входят в состав одной двигательной единицы (ДЕ). Скелетные мышцы человека состоят из ДЕ всех трех типов. Одни из них включают преимущественно медленные ДЕ, другие — быстрые, третьи — и те, и другие.
Критерий разделения I типа IIa типа IIb типа Скорость сокращения
(определяется по миозиновой АТФазе).Медленные (частота нервных импульсов до 25 Гц) Средняя (25-50 Гц) Быстрые (частота нервных импульсов 50-100 Гц) Обмен веществ
(определяется по ферментам аэробных процессов, по ферментам митохондрий: сукцинатдегидрогеназе или СДГ)Окислительный (с кислородом) Смешанный Гликолитический (без кислорода) Цвет
(зависит от количества миоглобина)Красные (много миоглобина и митохондрий) Светло-красный (красный) Белые (мало миоглобина и митохондрий) Порог активации Низкий Средний Высокий Диаметр 50 мкм 80 мкм 100 мкм Утомление (при постоянной нагрузке) Снижение силы на 50% через несколько часов Снижение силы на 50% через 10 мин Снижение силы на 50% через 1,5 мин Быстрые и медленные мышечные волокна [ править | править код ]

Классифицируются по активности фермента миозиновой АТФ-азы и, соответственно, по скорости сокращения мышц. Волокна, содержащие миозин с высокой активностью АТФазы, относят к быстрым волокнам, а те, что содержат миозин с более низкой активностью АТФазы, — к медленным.

Активность АТФазы наследуется и тренировки не влияют на соотношение быстрых и медленных волокон. Освобождение энергии, заключенной в АТФ, осуществляется благодаря АТФ-азе. Энергии одной молекулы АТФ достаточно для одного поворота (гребка) миозиновых мостиков. Мостики расцепляются с актиновым филаментом, возвращаются в исходное положение, сцепляются с новым участком актина и делают гребок. Скорость одиночного гребка одинакова у всех мышц. Для очередного гребка требуется новая молекула АТФ. В волокнах с высокой АТФ-азной активностью расщепление АТФ происходит быстрее, и за единицу времени происходит большее количество гребков мостиками, то есть мышца сокращается быстрее и, соответственно, сильнее.
Медленные окислительные волокна содержат множество митохондрий и обладают высокой способностью к окислительному фосфорилированию. Эти волокна могут содержать значительное количество липидов, но меньшее количество гликогена. Большая часть АТФ, произведенного такими волокнами, зависит от снабжения крови кислородом и топливных молекул. Эти волокна окружают многочисленные капилляры. Они также содержат большое количество связывающего кислород миоглобина, который увеличивает поглощение кислорода тканями и способствует небольшому внутриклеточному накоплению кислорода. Миоглобин придает темно-красный цвет, поэтому окислительные волокна часто называют красными мышечными волокнами.
В быстрых волокнах, также названных гликолитическими волокнами, напротив, содержится мало митохондрий, но они обладают высокой концентрацией гликолитических ферментов и большим запасом гликогена. Из-за ограниченного использования кислорода их окружает относительно небольшое количество капилляров, и они содержат мало миоглобина. Их называют белыми мышечными волокнами вследствие их более светлого цвета по сравнению с красными окислительными волокнами.
Гликолитические, промежуточные и окислительные волокна [ править | править код ]
Гликолитические волокна, как правило, намного больше в диаметре, чем окислительные волокна. Чем больше диаметр, тем больше максимальное растяжение, которого они могут достичь (т.е. тем они сильнее).
Читайте также: Виниловая ткань для проектора
Классифицируются по окислительному потенциалу мышцы, то есть по количеству митохондрий в мышечном волокне. Митохондрии – это клеточные органеллы, в которых глюкоза или жир расщепляется до углекислого газа и воды, ресинтезируя АТФ, необходимую для ресинтеза креатинфосфата. Креатинфосфат используется для ресинтеза миофибриллярных молекул АТФ, которые используются для мышечного сокращения. Вне митохондрий в мышцах также может происходить расщепление глюкозы до пирувата с ресинтезом АТФ, но при этом образуется молочная кислота, которая закисляет мышцу и вызывает ее утомление.
По этому признаку мышечные волокна подразделяются на три группы:
- Окислительные мышечные волокна. В них масса митохондрий так велика, что существенной прибавки ее в ходе тренировочного процесса уже не происходит.
- Промежуточные мышечные волокна. В них масса митохондрий значительно снижена, и в мышце в процессе работы накапливается молочная кислота, однако достаточно медленно, и утомляются они гораздо медленнее, чем гликолитические.
- Гликолитические мышечные волокна имеют очень незначительное количество митохондрий. Поэтому в них преобладает анаэробный гликолиз с накоплением молочной кислоты, отчего они и получили свое название.
У не тренирующихся людей обычно быстрые волокна — гликолитические и промежуточные, а медленные – окислительные. Однако при правильных тренировках на увеличение выносливости быстрые мышечные волокна превращаются из гликолитических в промежуточные. Также возможен переход промежуточных волокон в окислительные. При силовых тренировках промежуточные волокна могут переходить в гликолитические. При этом соотношение медленных и быстрых волокон генетически предопределено практически не меняется вне зависимости от тренировок (переход не более 1-3%).
- анаэробный гликолиз – расщепление глюкозы без кислорода до молочной кислоты с ресинтезом АТФ;
- аэробный гликолиз, или окисление, – расщепление пирувата в митохондриях с участием кислорода до углекислого газа, воды и ресинтезом АТФ.
Свойства различных типов мышечных волокон. Для классификации мышечных волокон в тексте использована система 1, но также приведены и названия, используемые в других системах
Медленные окислительные (МО) волокна
Быстрые окислительно-гликолитические (БОГ) волокна
Быстрые гликолитические (БГ) волокна
Основной источник образования АТФ
Тип миозиновой АТФ-азной активности
Активность гликолитических ферментов
Скорость наступления усталости
Двигательная единица — это один мотонейрон и иннервируемые им мышечные волокна. Когда МО мотонейрон стимулирует свои волокна, сокращается гораздо меньше мышечных волокон, чем когда свои волокна стимулирует мотонейрон БГ. Следовательно, двигательные БГ волокна достигают пикового напряжения быстрее и, взятые вместе, развивают большее усилие, чем МО волокна.
Волокна скелетных мышц различаются также по их способности противостоять усталости. Утомление БГ волокон происходит быстрее, тогда как МО волокна очень устойчивы к усталости. Быстро окисляющиеся волокна обладают промежуточной способностью сопротивляться усталости. Характеристики различных типов волокон скелетных мышц отображены в табл. 1.
Все мышцы человека обладают разным процентным соотношением БГ и МО мышечных волокон. В зависимости от доли имеющихся типов волокон, мышцы могут значительно различаться по максимальной скорости сокращения, силе и утомляемости. Например, в икроножных мышцах наблюдается преобладание БГ волокон, придающее им способность к сильному и быстрому сокращению, которое используется, например, при прыжках. С другой стороны, в камбаловидной мышце больше МО мышечных волокон, и она используется при длительной активности мышц ног.
В целом, МО мышечные волокна обладают высоким уровнем аэробной выносливости. Способность поддерживать мышечную активность в течение длительного времени известна как мышечная выносливость. Так как МО волокна обладают высокой аэробной выносливостью, они чаще всего задействуются во время нагрузок на выносливость (например, в марафонском беге) и во время большинства повседневных занятий, где требования к мышечной силе невысоки (например, ходьба).
БГ мышечные волокна, с другой стороны, обладают относительно низкой аэробной выносливостью. При нормальной, малоинтенсивной деятельности БГ волокна используются довольно редко, но при «взрывных» нагрузках они преобладают. Предполагается, что они активизируются, когда во время физической нагрузки оказывается превышен анаэробный порог; тогда уровень молочной кислоты в крови и в мышечных волокнах начинает повышаться немного раньше.
БОГ двигательные единицы генерируют гораздо большую силу, чем МО двигательные единицы, но они легко устают из-за своей ограниченной выносливости. Поэтому БОГ волокна, по всей видимости, используются в основном при непродолжительной интенсивной нагрузке на выносливость, например при пробежке на 1 милю или заплыве на 400 м.
Как было доказано, спортивные тренировки не меняют относительное соотношение БГ и МО волокон. По всей видимости, напротив, — это почти полностью определяется генетическим наследованием, и это, в свою очередь, может определять основные спортивные способности разных людей. С практической точки зрения двигательный нерв определяет тип мышечных волокон в двигательной единице. Если нерв, иннервирующий медленную двигательную единицу, отрезать и соединить с другим нервным волокном, иннервирующим быструю двигательную единицу, эта прежде быстрая двигательная единица может постепенно измениться и стать медленной. Варьирующиеся доли волокон разных типов в четырехглавых мышцах некоторых спортсменов, представляющих различные виды спорта, показаны в табл. 2.
Многие люди интересуются конными скачками. Лошади также участвуют в Олимпийских играх
естественно, не одни. Финская лошадь (ее мышцы примерно на 70% состоят из быстрых волокон) может бежать со скоростью 12,5 м/с, в то время как лошадь для состязаний на короткие дистанции (быстрые волокна составляют около 90% ее мышц) может развить скорость 20 м/с — это явный признак различия их свойств, которые развивались в течение многих поколений.
Таблица 2. Процентное соотношение МО и БГ волокон в четырехглавых мышцах спортсменов по сравнению с обычным человеком
Медленные окислительные волокна (%)
Быстрые гликолитические волокна (%)
Бегуны на марафонскую дистанцию
В мышечном веретене мышц также содержатся совершенно разные типы мышечных волокон. Эти структуры воспринимают напряжение мышц. Чувствительность мышечных веретен может быть отрегулирована при сокращении их особых интрафузальных мышечных волокон. Веретена расположены параллельно основной мышце или экстра-фузальным волокнам. Уровнем сокращения интрафузальных мышечных волокон в веретенах управляют гамма-мотонейроны, тогда как альфа-мотонейроны регулируют экстра-фузальные мышечные волокна, которые непосредственно отвечают за сокращение мышц.
Высокопороговые и низкопороговые волокна [ править | править код ]
Классифицируются по уровню порога возбудимости двигательных единиц. Мышца сокращается под действием нервных импульсов, которые имеют электрическую природу. Каждая двигательная единица (ДЕ) включает в себя мотонейрон, аксон и совокупность мышечных волокон. Количество ДЕ у человека остается неизменным на протяжении всей жизни. Двигательные единицы имеют свой порог возбудимости. Если нервные импульсы, посылаемые мозгом, имеют частоту ниже этого порога, ДЕ пассивна. Если нервные импульсы имеют пороговую для этой ДЕ величину или превышают ее, мышечные волокна активируются и начинают сокращаться. Низкопороговые ДЕ имеют маленькие мотонейроны, тонкий аксон и сотни иннервируемых медленных мышечных волокон. Высокопороговые ДЕ имеют крупные мотонейроны, толстый аксон и тысячи иннервируемых быстрых мышечных волокон.
Медленные окислительные волокна относятся к низкопороговым (возбуждаются при незначительной нагрузке). Быстрые волокна относятся к высокопороговым (включатся только при интенсивной нагрузке).
Эндокринология и мышечные волокна [ править | править код ]
Существование различных типов мышечных волокон обеспечивает значительную гетерогенность тканей скелетных мышц и их способность выполнять разнообразные функциональные задачи. Иммуногистохимический и биохимический анализ скелетных мышц показал, что такое структурно-функциональное разнообразие мышечных волокон обусловлено существованием широкого спектра изоформ миозина. Миозин — молекула, от которой наряду с актином зависит мышечное сокращение. Молекула миозина состоит из двух тяжелых цепей (МуНС) и четырех легких цепей (MyLC) (Schiaffino, Reggiani, 1996; Pette, Staron, 1997). Тяжелые цепи миозина представлены несколькими изоформами, от свойств которых зависят скоростно-силовые качества мышечных волокон.
Читайте также: Кигуруми какая ткань лучше
В скелетных мышцах взрослого человека происходит экспрессия четырех наиболее важных изоформ МуНС: MyHCip, MyHCIIA, MyHCIIX/IID и МуНСПВ. Каждая изоформа характеризуется специфической скоростью сокращения и развиваемым усилием. Волокна, содержащие MyHCI, отличаются низкой скоростью сокращения и развивают меньшее усилие по сравнению с волокнами, содержащими MyHCIIA, ИХ и IIB. Среди волокон, состоящих из быстрых МуНС, наиболее быстрыми и сильными являются те, которые построены из МуНСПВ, за ними следуют волокна, в состав которых входят МуНСИХ и MyHCIIA (Bottineli et al., 1994a, 1994b).
Занятия физическими упражнениями могут приводить к существенным изменениям сократительных свойств скелетных мышц. Принято считать, что тренировка выносливости сопровождается увеличением количества медленных изоформ миозина (Baumann et al., 1987; Schaub et al., 1989). В то же время силовая тренировка вызывает увеличение MyHCIIA и уменьшение МуНСПХ (Staron et al., 1991; Adams et al., 1993; Andersen J.L. et al., 1994; Fry et al., 1994; Kraemer et al., 1995; Kadi, Thorncll, 1999; Andersen J.L., Aagaard, 2000). Кроме того, предполагается, что мышечные волокна, содержащие МуНСИХ, у основной массы людей очень редко вовлекаются в выполнение работы в процессе обычной ежедневной активности. Если они начинают вовлекаться в выполнение работы, например в процессе физической тренировки, то превращаются в волокна, содержащие MyHCIIA (волокна, включающие эту изоформу тяжелых цепей миозина, обладают большей выносливостью по сравнению с волокнами типа ИХ) (Goldspink G. et al., 1991; Staron et al., 1991; Kraemer et al., 1995). Во время тренировки мышечной силы или выносливости происходит значительное изменение гормонального фона скелетных мышц, которое является мощным сигналом, способным запустить процесс изменения содержания изоформ миозина в мышцах, подвергающихся физической нагрузке.
Влияние тестостерона [ править | править код ]
В некоторых экспериментах на животных после применения андрогенных анаболических стероидов наблюдали изменение соотношения изоформ тяжелых цепей миозина в сторону увеличения медленных изоформ (Fritzshe et al., 1994; Czesla ct al., 1997). Сообщалось об увеличении доли волокон, содержащих MyHCIIA, наряду с сокращением количества волокон, содержащих МуНСПВ, в ряде скелетных мышц грызунов после применения андрогенных анаболических стероидов (Eggington, 1987; Dimauro et al., 1992). Однако сообщалось также о том, что андрогенные стероиды вызывают уменьшение доли мышечных волокон, содержащих MyHCIIA, по отношению к волокнам, состоящим из МуНСПВ (Kelly et al., 1985; Lyons et al., 1986; Salmons, 1992). Эти результаты говорят о том, что характер воздействия андрогенных анаболических стероидов на сократительные способности может зависеть от типа мышц и у различных видов может быть разным. Действительно, существуют и другие данные, свидетельствующие об отсутствии какого-либо воздействия андрогенных анаболических стероидов по соотношение мышечных волокон, содержащих различные изоформы МуНС. Например, в экспериментах на животных чрезмерная нагрузка мышц вызывала увеличение содержания медленных MyHCI, и дополнительное использование андрогенных анаболических стероидов не влияло на характер содержания тяжелых цепей миозина (Boissonneault et al., 1987). Точно так прием андрогенных анаболических стероидон не вызывал изменений сдвига соотношения изоформ МуНС, вызванного экспериментами с обездвиживанием нижней конечности (Tsika et al., 1987). Наконец, не удалось обнаружить никаких различий в соотношении разных изоформ МуНС в трапециевидной мышце хорошо тренированных тяжелоатлетов, принимавших и не принимавших андрогенные анаболические стероиды (Kadi et al., 1999b).
Влияние эстрогенов [ править | править код ]
Хорошо известен тот факт, что уменьшение развиваемой силы происходит в менопаузе (Greeves et al., 1999; Dionne et al., 2000; Meeuwsen et al., 2000). Ha клеточном уровне показано, что удаление яичников сопровождается изменением соотношения изоформ тяжелых цепей миозина в сторону увеличения медленных волокон и понижением спонтанного бега у крыс (Kadi et al., 2000). В целом изменения соотношения изоформ МуНС имеют следующую тенденцию: МуНС I Влияние соматотропного гормона [ править | править код ]
Сообщается о том, что прием соматотропного гормона (СТГ) индуцирует увеличение количества МуНСИХ в латеральной широкой мышце бедра у здоровых мужчин старшего возраста (Lange et al., 2002). Изменение соотношения изоформ МуНС в сторону увеличения МуНСИХ авторы исследования рассматривали как “омоложение” состава тяжелых цепей миозина, поскольку старение обычно сопровождается уменьшением доли МуНСИХ в этой группе мышц (Lange et al., 2002). Однако доля МуНСИХ у пациентов с дефицитом СТГ была выше по сравнению с основной массой здорового населения (Daugaard et al., 1999). Более того, после лечения больных с дефицитом СТГ препаратами рекомбинантного гормона роста в течение 6 месяцев у них не было выявлено никаких изменений в соотношении различных изоформ МуНС (Daugaard et al., 1999). Аналогичным образом было показано, что применение СТГ у крыс приводит к существенному увеличению поперечного сечения мышечных волокон типа II в камбаловидной мышце, не оказывая заметного влияния на содержание различных изоформ в составе мышечных волокон (Aroniadou-Anderjaska et al., 1996). Вопрос о том, приводит ли повышение уровня СТГ к изменению соотношения изоформ МуНС в сторону увеличения быстрых изоформ миозина, требует дальнейших исследований.
Влияние гормонов щитовидной железы [ править | править код ]
Гормоны щитовидной железы, или тироидные гормоны, оказывают сильное регуляторное воздействие на соотношение различных изоформ тяжелой цепи миозина в составе скелетной мышцы (D’Albis, Butler-Browne, 1993). Показано, что регуляция соотношения МуНС в скелетных мышцах крысы является специфической для пола и типа мышц (Larsson, Yu, 1997). Применение 3.5.3’-трийодтиронина (Т3) приводит к подавлению содержания MyHCI и увеличению содержания MyHCIIA в камбаловидной мышце самцов и самок, тогда как стимуляция содержания МуНСИХ наблюдалась только в мышцах самцов крыс (Larsson, Yu, 1997). Применение Т3 не вызывало никаких изменений в длинном разгибателе пальцев стопы у самцов крыс. В то же время в аналогичной ситуации в той же мышце у самок отмечалось достоверное изменение соотношения изоформ MyHCIIA и ИВ в пользу увеличения последней (Larsson, Yu, 1997). В целом эти результаты показывают, что сократительные качества скелетных мышц находятся под контролем ряда гормонов и ростовых факторов и изменение гормонального фона в этих мышцах при выполнении физических упражнений может быть в определенной степени ответственным за изменение характеристик мышцы в соответствии с физиологическими потребностями. Становится все более очевидным, что изменения структуры и функции мышц, происходящие под влиянием изменений гормонального фона, могут зависеть от пола и типа мышц.
Заключение [ править | править код ]
Были рассмотрены лишь отдельные аспекты значения специфических гормонов и ростовых факторов в регуляции некоторых важных параметров мышц, определяющих спортивные показатели. Эта сфера мышечной физиологии только начинает развиваться и здесь еще многое предстоит открыть прежде чем станет понятно взаимоотношение различных факторов, принимающих участие в разнообразных процессах адаптации скелетных мышц к различным видам двигательной активности. Последовательное описание различных этапов адаптации мышц к двигательной активности позволит создать основу для концепции индивидуализированного выбора упражнений с целью оптимизации качества тренировочных программ как для хорошо физически подготовленных лиц, так и для тех, кто ведет малоподвижный образ жизни, а также для специальных групп населения.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом