
Цельная кровь состоит из жидкой части (плазмы) и форменных элементов, к которым относят эритроциты, лейкоциты и кровяные пластинки — тромбоциты.
Функции крови:
1) транспортная — перенос газов (02 и С02), пластических (аминокислот, нуклеозидов, витаминов, минеральных веществ), энергетических (глюкоза, жиры) ресурсов к тканям, а конечных продуктов обмена — к органам выделения (желудочно-кишечный тракт, легкие, почки, потовые железы, кожа);
2) гомеостатическая — поддержание температуры тела, кислотно-основного состояния организма, водно-солевого обмена, тканевого гомеостаза и регенерации тканей;
3) защитная — обеспечение иммунных реакций, кровяного и тканевого барьеров против инфекции;
4) регуляторная — гуморальной и гормональной регуляции функций различньгх систем и тканей;
5) секреторная — образование клетками крови биологически активных веществ.
Функции и свойства эритроцитов
Эритроциты переносят 02 содержащимся в них гемоглобином от легких к тканям и С02 от тканей к альвеолам легких. Функции эритроцитов обусловлены высоким содержанием гемоглобина (95 % массы эритроцита), деформируемостью цитоскелета, благодаря чему эритроциты легко проникают через капилляры с диаметром меньше 3 мкм, хотя имеют диаметр от 7 до 8 мкм. Глюкоза является основным источником энергии в эритроците. Восстановление формы деформированного в капилляре эритроцита, активный мембранный транспорт катионов через мембрану эритроцита, синтез глютатиона обеспечиваются за счет энергии анаэробного гликолиза в цикле Эмбдена—Мейергофа. В ходе метаболизма глюкозы, протекающего в эритроците по побочному пути гликолиза, контролируемого ферментом дифосфоглицератмутазой, в эритроците образуется 2,3-дифосфоглицерат (2,3-ДФГ). Основное значение 2,3-ДФГ заключается в уменьшении сродства гемоглобина к кислороду.
В цикле Эмбдена—Мейергофа расходуется 90 % потребляемой эритроцитами глюкозы. Торможение гликолиза, возникающее, например, при старении эритроцита и уменьшающее в эритроците концентрацию АТФ, приводит к накоплению в ней ионов натрия и воды, ионов кальция, повреждению мембраны, что понижает механическую и осмотическую устойчивость эритроцита, и стареющий эритроцит разрушается. Энергия глюкозы в эритроците используется также в реакциях восстановления, защищающих компоненты эритроцита от окислительной денатурации, которая нарушает их функцию. Благодаря реакциям восстановления атомы железа гемоглобина поддерживаются в восстановленной, т. е. двухвалентной форме, что препятствует превращению гемоглобина в метгемоглобин, в котором железо окислено до трехвалентного, вследствие чего метгемоглобин неспособен к транспорту кислорода. Восстановление окисленного железа метгемоглобина до двухвалентного обеспечивается ферментом — метгемоглобинредуктазой. В восстановленном состоянии поддерживаются и серусодержащие группы, входящие в мембрану эритроцита, гемоглобин, ферменты, что сохраняет функциональные свойства этих структур.
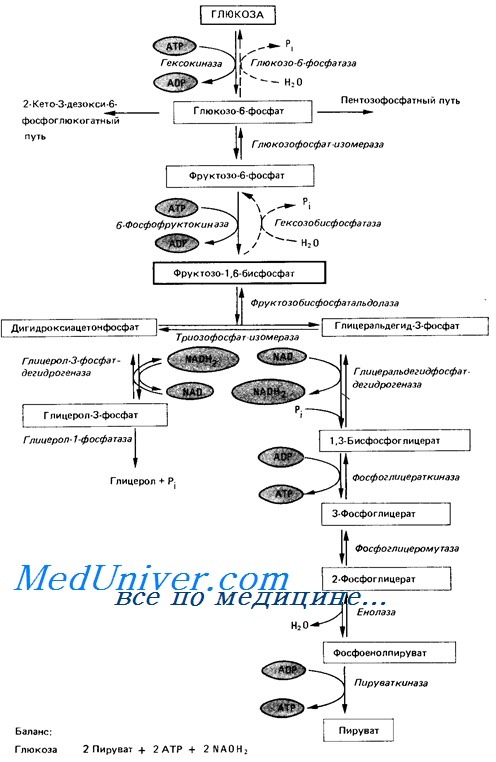
Цикл Эмбден-Мейергоффа эритроцитов
Эритроциты имеют дисковидную, двояковогнутую форму, их поверхность — около 145 мкм2, а объем достигает 85—90 мкм3. Такое соотношение площади к объему способствует деформабильно-сти (под последней понимают способность эритроцитов к обратимым изменениям размеров и формы) эритроцитов при их прохождении через капилляры. Форма и деформабильность эритроцитов поддерживаются липидами мембран — фосфолипидами (глицерофосфолипидами, сфинголипидами, фосфотидилэтаноламином, фосфатидилсирином и др.), гликолипидами и холестерином, а также белками их цитоскелета. В состав цитоскелета мембраны эритроцита входят белки — спектрин (основной белок цитоскелета), анкирин, актин, белки полосы 4.1, 4.2, 4.9, тропомиозин, тропомодулин, адцуцин. Основой мембраны эритроцита является липидный бислой, пронизанный интегральными белками цитоскелета — гликопротеинами и белком полосы 3. Последние связаны с частью белковой сети цитоскелета — комплексом спектрин—актин—белок полосы 4.1, локализованным на цитоплазматической поверхности липидного бислоя мембраны эритроцита (рис. 7.1).
Взаимодействие белкового цитоскелета с липидным бислоем мембраны обеспечивает стабильность структуры эритроцита, поведение эритроцита как упругого твердого тела при его деформации. Нековалентные межмолекулярные взаимодействия белков цитоскелета легко обеспечивают изменение размеров и формы эритроцитов (их деформацию) при прохождении этих клеток через микроциркуляторное русло, при выходе ретикулоцитов из костного мозга в кровь — благодаря изменению расположения молекул спектрина на внутренней поверхности липидного бислоя. Генетические аномалии белков цитоскелета у человека сопровождаются появлением дефектов мембраны эритроцитов. В результате последние приобретают измененную форму (так называемые сфероциты, элиптоциты и др.) и имеют повышенную склонность к гемолизу. Увеличение соотношения холестерин—фосфолипиды в мембране увеличивает ее вязкость, уменьшает текучесть и эластичность мембраны эритроцита. В результате снижается деформируемость эритроцита. Усиление окисления ненасыщенных жирных кислот фосфолипидов мембраны перекисью водорода или супероксидными радикалами вызывает гемолиз эритроцитов (разрушение эритроцитов с выходом гемоглобина в окружающую среду), повреждение молекулы гемоглобина эритроцита. Постоянно образующийся в эритроците глютатион, а также антиоксиданты (остокоферол), ферменты — глутатионредуктаза, супероксиддисмутаза и др. защищают компоненты эритроцита от этого повреждения.
Читайте также: Опорные защитные ткани у растений
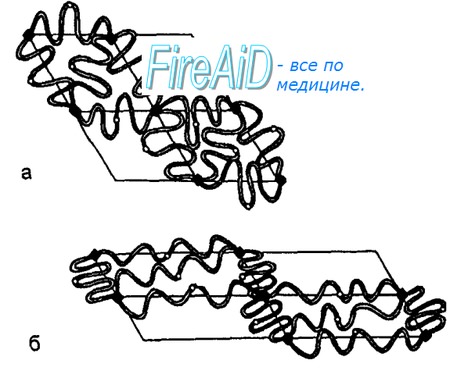
Рис. 7.1. Схема модели изменений цитоскелета мембраны эритроцита во время его обратимой деформации. Обратимая деформация эритроцита изменяет лишь пространственную конфигурацию (стереометрию) эритроцита, следующую за изменением пространственного расположения молекул цитоскелета. При этих изменениях формы эритроцита площадь поверхности эритроцита остается неизменной. а — положение молекул цитоскелета мембраны эритроцита при отсутствии его деформации. Молекулы спектрина находятся в свернутом состоянии.
До 52 % массы мембраны эритроцитов составляют белки гликопротеины, которые с олигосахаридами образуют антигены групп крови. Глико-протеины мембраны содержат сиаловую кислоту, которая придает отрицательный заряд эритроцитам, отталкивающий их друг от друга.
Энзимы мембраны — Ка+/К+-зависимая АТФаза обеспечивает активный транспорт Na+ из эритроцита и К+ в его цитоплазму. Са2+-зависимая АТФаза выводит Са2+ из эритроцита. Фермент эритроцита карбоангидраза катализирует реакцию: Са2+ Н20 Н2С03 о Н+ + НСО3, поэтому эритроцит транспортирует часть углекислого газа от тканей к легким в виде бикарбоната, до 30 % С02 переносится гемоглобином эритроцитов в форме карбаминового соединения с радикалом NH2 глобина.
Эритроциты
Общий миелоидный прародитель → Проэритробласт → Мегалобласт → Полихроматический эритробласт → Нормоцит → Ретикулоцит → Эритроцит
В данной статье или разделе имеется список источников или внешних ссылок, но источники отдельных утверждений остаются неясными из-за отсутствия сносок.
Эритроци́ты (от греч. ἐρυθρός — красный и κύτος — вместилище, клетка), также известные под названием кра́сные кровяны́е тельца́, — клетки крови человека, позвоночных животных и некоторых беспозвоночных (сипункулид, у которых эритроциты плавают в полости целома [1] ).
Функции
Эритроциты — высокоспециализированные клетки, функцией которых является перенос кислорода из лёгких к тканям тела и транспорт диоксида углерода (CO2) в обратном направлении. У позвоночных, кроме млекопитающих, эритроциты имеют ядро, у эритроцитов млекопитающих ядро отсутствует.
Наиболее специализированы эритроциты млекопитающих, лишённые в зрелом состоянии ядра и органелл и имеющие форму двояковогнутого диска, обуславливающую высокое отношение площади к объёму, что облегчает газообмен. Особенности цитоскелета и клеточной мембраны позволяют эритроцитам претерпевать значительные деформации и восстанавливать форму (эритроциты человека диаметром 8 мкм проходят через капилляры диаметром 2—3 мкм).
Транспорт кислорода обеспечивается гемоглобином (Hb), на долю которого приходится ≈98 % массы белков цитоплазмы эритроцитов (в отсутствии других структурных компонентов). Гемоглобин является тетрамером, в котором каждая белковая цепь несёт гем — комплекс протопорфирина IX с ионом двухвалентного железа, кислород обратимо кординируется с ионом Fe 2+ гемоглобина, образуя оксигемоглобин HbO2:

Hb + O2 HbO2
Особенностью связывания кислорода гемоглобином является его аллостерическое регулирование — стабильность оксигемоглобина падает в присутствии 2,3-дифосфоглицериновой кислоты — промежуточного продукта гликолиза и, в меньшей степени, углекислого газа, что способствует высвобождению кислорода в тканях, в нём нуждающихся.
Транспорт углекислого газа эритроцитами происходит с участием карбоангидразы, содержащейся в их цитоплазме. Этот фермент катализирует обратимое образование бикарбоната из воды и углекислого газа, диффундирующего в эритроциты:
H2O + CO2 H + + HCO3 –
В результате в цитоплазме накапливаются ионы водорода, однако снижение pH при этом незначительно из-за высокой буферной ёмкости гемоглобина. Вследствие накопления в цитоплазме ионов бикарбоната возникает градиент концентрации, однако ионы бикарбоната могут покидать клетку только при условии сохранения равновесного распределения зарядов между внутренней и внешней средой, разделённых цитоплазматической мембраной, то есть выход из эритроцита иона бикарбоната должен сопровождаться либо выходом катиона, либо входом аниона. Мембрана эритроцита практически непроницаема для катионов, но содержит хлоридные ионые каналы, в результате выход бикарбоната из эритроцита сопровождается входом в него хлорида (хлоридный сдвиг).
Формирование эритроцитов
Формирование эритроцитов (эритропоэз) происходит в костном мозге черепа, рёбер и позвоночника, а у детей — ещё и в костном мозге в окончаниях длинных костей рук и ног. Продолжительность жизни — 3—4 месяца, разрушение (гемолиз) происходит в печени и селезёнке. Прежде чем выйти в кровь, эритроциты последовательно проходят несколько стадий пролиферации и дифференцировки в составе эритрона — красного ростка кроветворения.
Полипотентная стволовая клетка крови (СКК) даёт клетку-предшественницу миелопоэза (КОЕ-ГЭММ), которая в случае эритропоэза даёт клетку-родоначальницу миелопоэза (КОЕ-ГЭ), которая уже даёт унипотентную клетку, чувствительную к эритропоэтину (БОЕ-Э).
Бурстобразующая единица эритроцитов (БОЕ-Э) даёт начало эритробласту, который через образование пронормобластов уже дают морфологически различимые клетки-потомки нормобласты (последовательно переходящие стадии):
- базофильные нормобласты (имеют базофильное ядро и цитоплазму, начинает синтезироваться гемоглобин),
- полихроматофильные нормобласты (ядро становится меньше, участки с гемоглобином приобретают оксифильность),
- оксифильные нормобласты (их ядро расположено на одном конце уже овальной клетки, не способны к делению, содержат много гемоглобина),
- ретикулоциты (безъядерные, содержат остатки органелл, главным образом шероховатой эндоплазматической сети). Ретикулоциты далее становятся эритроцитами.
Гемопоэз (в данном случае эритропоэз) исследуется по методу селезёночных колоний.
Большая клетка с ядром, не обладающая характерным красным цветом, — мегалобласт; затем она окрашивается в красный цвет — теперь это эритробласт. Нормоцит (нормобласт) уменьшается в размере в процессе развития. После утраты ядра нормоцит превращается в ретикулоцит.
У птиц, пресмыкающихся, земноводных и рыб ядро просто теряет активность, но сохраняет способность реактивации. Одновременно с исчезновением ядра по мере взросления эритроцита из его цитоплазмы исчезают рибосомы и другие компоненты, участвующие в синтезе белка. Ретикулоциты попадают в кровеносную систему и через несколько часов становятся полноценными эритроцитами.
Структура и состав
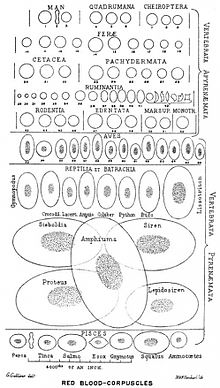

У большинства групп позвоночных эритроциты имеют ядро и другие органоиды.
У млекопитающих зрелые эритроциты лишены ядер, внутренних мембран и большинства органоидов. Ядра выбрасываются из клеток-предшественников в ходе эритропоэза. Обычно эритроциты млекопитающих имеют форму двояковогнутого диска и содержат в основном дыхательный пигмент гемоглобин. У некоторых животных (например, верблюда) эритроциты имеют овальную форму.
Содержимое эритроцита представлено главным образом дыхательным пигментом гемоглобином, обусловливающим красный цвет крови. Однако на ранних стадиях количество гемоглобина в них мало, и на стадии эритробластов цвет клетки синий; позже клетка становится серой и, лишь полностью созрев, приобретает красную окраску.

Важную роль в эритроците выполняет клеточная (плазматическая) мембрана, пропускающая газы (кислород, углекислый газ), ионы (Na, K) и воду. Плазмолемму пронизывают трансмембранные белки — гликофорины, которые, благодаря большому количеству остатков сиаловой кислоты, ответственны примерно за 60 % отрицательного заряда на поверхности эритроцитов.
На поверхности липопротеидной мембраны находятся специфические антигены гликопротеидной природы — агглютиногены — факторы систем групп крови (на данный момент изучено более 15 систем групп крови: AB0, резус фактор, антиген Даффи (англ.) русск. , антиген Келл, антиген Кидд (англ.) русск. ), обусловливающие агглютинацию эритроцитов при действии специфических агглютининов.
Эффективность функционирования гемоглобина зависит от величины поверхности соприкосновения эритроцита со средой. Суммарная поверхность всех эритроцитов крови в организме тем больше, чем меньше их размеры. У низших позвоночных эритроциты крупные (например, у хвостатого земноводного амфиумы — 70 мкм в диаметре), эритроциты высших позвоночных мельче (например, у козы — 4 мкм в диаметре). У человека диаметр эритроцита составляет 7,2—7,5 мкм, толщина — 2 мкм, объём — 76-110 мкм³ [источник не указан 1292 дня] .
В одном литре крови содержится эритроцитов:
- у мужчин 4,5·10 12 /л—5,5·10 12 /л (4,5—5,5 млн в 1 мм³ крови),
- у женщин — 3,7·10 12 /л—4,7·10 12 /л (3,7—4,7 млн в 1 мм³),
- у новорождённых — до 6,0·10 12 /л (до 6 млн в 1 мм³),
- у пожилых людей — 4,0·10 12 /л (меньше 4 млн в 1 мм³).
Переливание крови
При переливании крови от донора к реципиенту возможна агглютинация (склеивание) и гемолиз (разрушение) эритроцитов. Чтобы этого не происходило, необходимо учитывать группы крови, открытые К. Ландштейнером и Я. Янским в 1900 году. Агглютинацию вызывают белки, находящиеся на поверхности эритроцита — антигены (агглютиногены) и находящиеся в плазме антитела (агглютинины). Существуют 4 группы крови, для каждой характерны различные антигены и антитела. Переливание обычно проводится лишь между обладателями одной группы крови.
| I — 0 | II — A | III — B | IV — AB |
|---|---|---|---|
| αβ | β | α | — |
Место в организме
Форма двояковогнутого диска обеспечивает прохождение эритроцитов через узкие просветы капилляров. В капиллярах они движутся со скоростью 2 сантиметра в минуту, что даёт им время передать кислород от гемоглобина к миоглобину. Миоглобин действует как посредник, принимая кислород у гемоглобина в крови и передавая его цитохромам в мышечных клетках.
Количество эритроцитов в крови в норме поддерживается на постоянном уровне (у человека в 1 мм³ крови 4,5—5 млн эритроцитов, у некоторых копытных 15,4 млн (лама) и 13 млн (коза) эритроцитов, у пресмыкающихся — от 500 тыс. до 1,65 млн, у хрящевых рыб — 90—130 тыс.) Общее число эритроцитов снижается при анемиях, повышается при полицитемии.
Продолжительность жизни эритроцита человека в среднем 125 суток (ежесекундно образуется около 2,5 млн эритроцитов и такое же их количество разрушается), у собак — 107 дней, у кроликов и кошек — 68.
Патология

При различных заболеваниях крови возможно изменение цвета эритроцитов, их размеров, количества, а также формы; они могут принимать, например, серповидную, овальную, сферическую или мишеневидную форму.
Изменение формы эритроцитов называется пойкилоцитозом (англ.) русск. . Сфероцитоз (сферическая форма эритроцитов) наблюдается при некоторых формах наследственной анемии. Эллиптоциты (эритроциты овальной формы) встречаются при мегалобластной и железодефицитной анемии, талассемиях и других заболеваниях. Акантоциты и эхиноциты (эритроциты шиповатой формы) встречаются при поражениях печени, наследственных дефектах пируваткиназы и др. Мишеневидные эритроциты (кодоциты) — это клетки с бледной тонкой периферией и центральным утолщением, содержащем скопление гемоглобина. Встречаются при талассемиях и других гемоглобинопатиях, интоксикации свинцом и др. Серповидные эритроциты — признак серповидноклеточной анемии. Встречаются и другие формы эритроцитов [2] .
При изменении кислотно-щелочного баланса крови в сторону закисления (от 7,43 до 7,33) происходит склеивание эритроцитов в виде монетных столбиков, либо их агрегация.
Среднее содержание гемоглобина для мужчин 13,3—18 г% (или 4,0—5,0·10 12 единиц), для женщин 11,7—15,8 г% (или 3,9—4,7·10 12 единиц). Единица измерения уровня гемоглобина представляет собой процент содержания гемоглобина в 1 грамме эритроцитарной массы.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом