
Соединительная ткань ретикулярная ткань лимфатического узла
Лимфатические узлы расположены по ходу лимфатических сосудов и вместе с ними составляют лимфатическую систему. Они являются органами лимфопоэза и образования антител. Лимфатические узлы, которые оказываются первыми на пути лимфатических сосудов, несущие лимфу из данной области тела (региона) или органа, считаются регионарными.
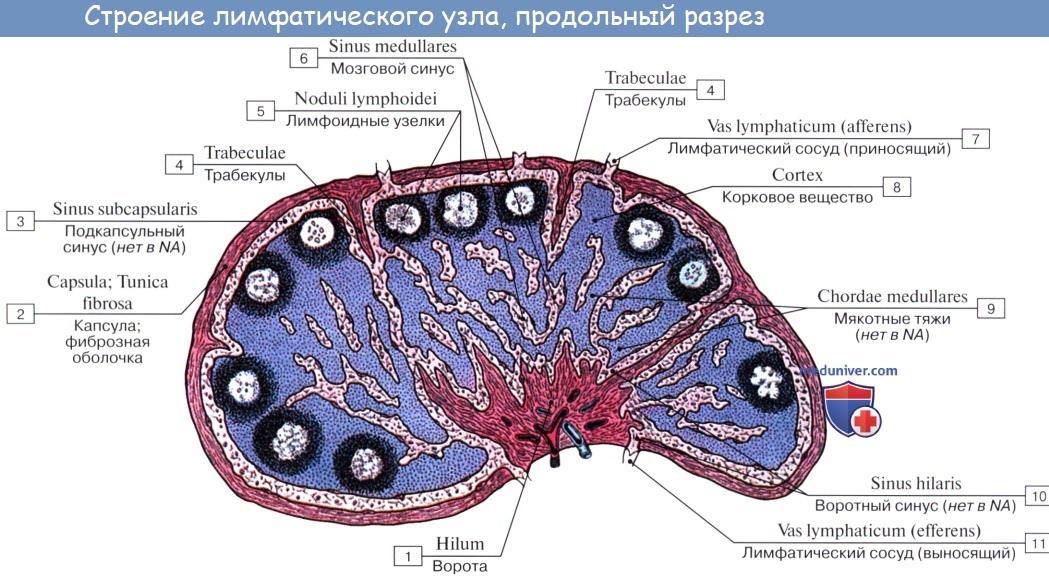
По описанию М. Р. Сапина, каждый лимфатический узел покрыт соединительнотканной капсулой (capsula nodi lymphatici), от которой внутрь узла отходят капсулярные трабекулы (trabeculae nodi lymphatici).
На поверхности узла имеется вдавление — ворота узла (hilus nodi lymphatici). У соматических узлов имеются одни ворота, у висцеральных встречается 3 — 4. Через ворота проникают в узел артерии и нервы, выходят вены и выносящие лимфатические сосуды. От капсулы в области ворот отходят в паренхиму узла воротные (хиларные) трабекулы. Воротные и капсулярные трабекулы соединяются, придавая лимфатическому узлу дольчатое строение.
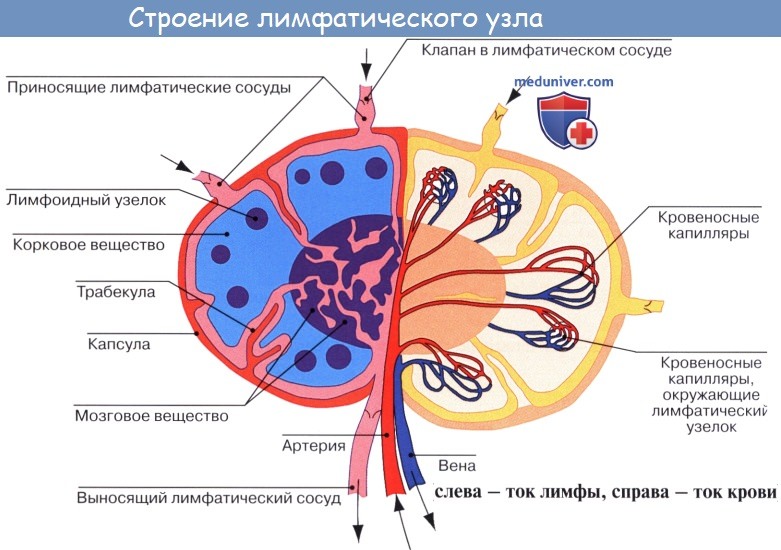
С капсулой узла и трабекулами связана строма узла, образованная ретикулярной соединительной тканью, в петлях которой находятся клетки крови, главным образом лимфоциты.
Ретикулярная ткань и лежащие в ее петлях клетки составляют паренхиму узла, которую подразделяют на корковое и мозговое вещество. В корковом веществе (близком к капсуле) располагаются мелкие узелки, или фолликулы (noduli s. folliculi lymphatici), содержащие преимущественно иммунокомпетент-ные клетки (В-лимфоциты). Мозговое вещество представлено мякотными тяжами (chorda medullaris), являющимися зоной скопления В-лимфоцитов, связанных с выработкой гуморального иммунитета.
Между капсулой, трабекулой и паренхимой имеются щели — лимфатические синусы (sinus nodi lymphatici). По синусам течет лимфа, поступившая в лимфатический узел. Она сначала поступает в краевой синус, находящийся под капсулой узла (sinus marginalis), 7 в который открываются приносящие лимфатические сосуды. Далее она проникает в синусы коркового и мозгового вещества, а затем в воротный синус (sinus hilaris) и из него в выносящие лимфатические сосуды. На своем пути лимфа как бы просачивается также через паренхиму узла и течет по краевому синусу более коротким путем от приносящих лимфатических сосудов к выносящим.
Сквозь стенки синусод; в паренхиму лимфатического узла проникают и там накапливаются инородные частицы, подвергающиеся ; воздействию лимфы.
Каждый лимфатический узел обильно кровоснабжается, причем артерии проникают в него не только через ворота, но и через капсулу.
Экспериментально доказан обмен в лимфатических узлах между кровью и лимфой (Ю. И. Бородин и сотр.).
Условно выделяют 3 типа лимфатических узлов:
Первый тип характеризуется, в частности, тем, что у него площадь коркового вещества несколько меньше площади мозгового. Лимфатические узлы первого типа быстро и интенсивно наполняются рентгеноконтрастной массой.
Лимфатические узлы второго типа компактные.Они характеризуются преобладанием массы коркового вещества над мозговым и рентгенологически медленным и слабым контрастированием. Транспортная функция таких узлов минимальна.
Чаще всего встречаются лимфатические узлы третьего типа — промежуточные. Масса коркового и мозгового вещества в них примерно одинакова. Рентгеноконтрастным веществом они заполняются хорошо. Их конструкция эффективно обеспечивает обработку лимфы и транспортную функцию.
Отмеченные вариации лимфатических узлов, индивидуальные особенности их конструкции и соответственно функциональные потенции в известной мере обусловливают различную выживаемость онкологических больных.
Лимфатические узлы перестраиваются в течение всей жизни, в том числе у пожилых и старых людей. От юношеского возраста (17 — 21 год) до пожилого (60 — 75 лет) количество их уменьшается в 1 1/2—2 раза. По мере увеличения возраста человека в узлах, преимущественно соматических, роисходят утолщение капсулы и трабекул, увеличение соединительной ткани, замещение паренхимы жировой тканью. Такие узлы теряют свои естественные строение и свойства, запустевают и становятся непроходимыми для лимфы. Число лимфатических узлов уменьшается и за счет срастания двух узлов, лежащих рядом, в более крупный лимфатический узел.
Читайте также: Клеточное строение хрящевой ткани
С возрастом меняется и форма узлов. В молодом возрасте преобладают узлы округлой и овальной формы, у пожилых и старых людей они как бы вытягиваются в длину.
Таким образом, у пожилых и старых людей количество функционирующих лимфатических узлов уменьшается за счет их атрофии и срастания друг с другом, в результате чего у лиц старшего возраста преобладают крупные лимфатические узлы.
РЕТИКУЛЯРНАЯ ТКАНЬ
Ретикулярная ткань (textus connectivus reticularis (LNH); лат. reticulum сетка; син. сетчатая ткань) — разновидность соединительной ткани, состоящей из ретикулярных клеток и ретикулярных волокон, заключенных в основное межклеточное вещество и образующих рыхлую трехмерную сеть, являющуюся основой кроветворных и лимфоидных органов.
Представления о Ретикулярной ткани формировались в течение многих десятилетий, но лишь в последние 10—15 лет, благодаря использованию достижений цитогенетики, радиобиологии, иммунологии, трансплантологии, были получены принципиально новые данные о ее дифференцировке, структуре и функции.
В эмбриогенезе Ретикулярной ткани дифференцируется из мезенхимы (см.) и в раннем постнатальном периоде постепенно приобретает строение зрелой ткани.

Ретикулярные клетки (cellulae reticulares) относят к клеткам фибробластического типа, наряду с фибробластами, хондробластами и остеобластами, объединяемыми под названием «механоциты». Ретикулярные клетки (рис. 1) имеют уплощенную, веретеновидную или звездчатую форму с гладкой поверхностью; ядро — угловатое или вытянутое. Степень развития органелл, в частности зернистой эндоплазматической сети и комплекса Гольджи (см. Гольджи комплекс), вариабельна и зависит от функционального состояния ретикулярной клетки. Соседние ретикулярные клетки или отростки одних и тех же клеток контактируют друг с другом посредством соединений типа промежуточных или десмосом (см.).
Ввиду того, что ретикулярные клетки часто трудно различить среди массы кроветворных клеток, их идентификация возможна только при использовании электронной микроскопии (см.), гистохимических методов исследования (см.) и методов иммуногистохимии (см. Иммуноморфология) или комбинации этих методов, позволяющих с высокой степенью достоверности выявлять ретикулярные клетки, а также идентифицировать их от внешне сходных с ними клеток системы мононуклеарных фагоцитов (см.).
Гистохимические свойства ретикулярных клеток определяются органными, а также видовыми особенностями. У человека ретикулярные клетки белой пульпы селезенки (см.) отличаются от макрофагов (см.) более низкой активностью эстеразы (см.) и кислой фосфатазы (см.), а от интердигитирующих клеток (разновидности мононуклеарных фагоцитов) — отсутствием активности АТФ-азы. В ретикулярных клетках лимфоцитарной короны лимф, фолликулов селезенки выявляется отчетливая активность 5′-нуклеотидазы (табл.).
Активность 5′-нуклеотидазы определяется также в ретикулярных клетках белой пульпы селезенки крыс и морских свинок, но отсутствует у кроликов. Ретикулярные клетки костного мозга мышей и крыс характеризуются активностью щелочной фосфатазы, локализующейся на их плазматической мембране. Этим признаком они отличаются от макрофагов, в к-рых выявляется активность кислой фосфатазы, локализующейся в лизосомах (см.). Такой морфол. тип ретикулярных клеток часто называют фибробластическим.
Читайте также: Тейпирование при ушибах мягких тканей

В светлых (герминативных) центрах лимфатических фолликулов селезенки и лимфатических узлов (см.) описан особый тип ретикулярных клеток — дендритные ретикулярные клетки. Их отличают крупные размеры, ядро неправильной формы с выраженным ядрышком, многочисленные гладкие пузырьки в цитоплазме. Характерным признаком дендритных ретикулярных клеток являются длинные ветвящиеся отростки с многочисленными складками, глубоко проникающие между окружающими лимфоидными клетками (рис. 2). Отростки дендритных ретикулярных клеток вместе с цитоплазматическими выпячиваниями иммунобластов (В-лимфоцитов) формируют сложную сеть в виде лабиринта. В межклеточном пространстве среди отростков локализуются микровезикулярные структуры и глобулярные плотные частицы диаметром 20—70 нм. В области контакта отростков дендритных ретикулярных клеток видны структуры типа десмосом (см.). В отличие от макрофагов с окрашивающимися включениями и других мононуклеарных фагоцитов дендритные ретикулярные клетки имеют более низкую активность эстеразы и кислой фосфатазы, но высокую активность 5′-нуклеотидазы. По ряду гистохимических признаков они могут быть сходны с мононуклеарными фагоцитами: макрофагами красной пульпы селезенки, металлофильными (аргирофильными) клетками краевой зоны лимф, фолликулов, макрофагами с окрашивающимися включениями. Решающим критерием для идентификации дендритных ретикулярных клеток является способность этих клеток связывать (но не фагоцитировать) иммунные комплексы на своей поверхности.
Ретикулярные волокна (fibrae reticulares), входящие в состав Р. т., состоят из фибрилл различного диаметра, заключенных в гомогенное, плотное основное межклеточное вещество. Фибриллы диаметром 20—50 нм имеют осевую исчерченность, характерную для зрелого коллагена. Тонкие фибриллы диаметром 10 нм локализуются по периферии ретикулярных волокон. Ретикулярные волокна всегда окутаны цитоплазмой ретикулярных клеток в виде чехла (инвагинированы в цитолемму), просвет к-рого всегда сообщается с межклеточным пространством. В лимфоидных органах сеть ретикулярных волокон развита сильнее, чем в костном мозге, причем в тимус-зависимых зонах лимфатических узлов она более рыхлая, чем в корковом и мозговом веществе.
По данным биохимического и иммунохимического анализов, ретикулярные волокна состоят из коллагена III типа (см. Коллаген) и неколлагенового компонента в виде аморфного межфибриллярного вещества с выраженными иммуногенными свойствами. В составе неколлагенового компонента идентифицированы белки (90%), углеводы (4%) и липиды (4%). Для выявления ретикулярных волокон широко используют различные варианты импрегнации серебром. В повседневную практику начинают входить иммуногистохимические и иммунофлюоресцентные методы выявления ретикулярных волокон (см. Иммуноморфология, Иммунофлюоресценция), в основе к-рых лежит использование специфических антисывороток к коллагенам разных типов.
Р. т. образует строму и является носителем специфических органных функций костного мозга и периферических лимфоидных органов (селезенки, лимф, узлов, солитарных и групповых лимф, фолликулов жел.-киш. тракта). После того как было экспериментально доказано, что ретикулярные клетки гистогенетически независимы от кроветворных, их стали относить к категории стромальных элементов, ответственных за создание специфического микроокружения, обеспечивающего миграцию, сортировку, репликацию и дифференцировку кроветворных и лимфоидных клеток. Ретикулярные клетки способны восстанавливать исходное микроокружение при повреждении органов, содержащих Р. т., или переносить его при эктопической трансплантации. Во всех случаях сначала восстанавливается ретикулярная строма, а затем она репопулируется (заселяется) кроветворными или лимфоидными клетками (см. Лимфоидная ткань). В костном мозге ретикулярные клетки формируют также адвентициальный слой синусоидных капилляров и в условиях нормального кроветворения покрывают до 60% их поверхности. Длинные ветвящиеся отростки ретикулярных клеток вступают в специфическое взаимодействие с кроветворными клетками гранулоцитарно-го ряда дифференцировки (см. Кроветворные органы). Концентрация ретикулярных клеток повышена вблизи эндоста.
Читайте также: В ателье ткани необходимо раскроить рулон
Экспериментальные данные свидетельствуют о сохранении в пост-натальном онтогенезе гистогенети-ческой близости между разными типами механоцитов (фибробластами, хрящевыми, костными, ретикулярными клетками). Так, при культивировании костного мозга in vitro возникают колонии-клоны фибробластов; нек-рые из этих колоний при обратной трансплантации в организм формируют костномозговой орган (участок костной ткани, окружающей костный мозг), заселяемый кроветворными клетками. Эти данные показывают, что среди ретикулярных клеток костного мозга имеются элементы, способные трансформироваться в истинные фибробласты (судя по их способности синтезировать коллаген I и III типов), и в то же время, проявляющие остеогенные свойства. В другой экспериментальной модели костномозговой орган возникает в результате последовательных морфогенетических процессов под влиянием индуктора — деминерализованного костного матрикса, имплантированного в подкожную соединительную ткань. По мнению Редди, Гея, Гея, Миллера (А. N. Beddi, В. Gay, S. Gay. E. J. Miller, 1977), в этом случае под влиянием индуктора происходит последовательная трансформация фибробластов в хрящевые, костные, а затем ретикулярные (стромальные) клетки. По-видимому, ретикулярные клетки костного мозга способны также трансформироваться в жировые клетки, участвующие в создании кроветворного микроокружения (в красном костном мозге). В норме ретикулярные клетки отличает высокая радиорезистентность, и они практически не делятся.
Существует мнение, что на поверхности дендритных ретикулярных клеток имеются мембранные рецепторы для иммуноглобулинов (см.), с к-рыми связываются иммунные комплексы антигенов с антителами. Согласно другому мнению, антигены (см.) просто медленно фильтруются через лабиринт, создаваемый отростками дендритных ретикулярных клеток. Связывание антигена происходит быстрее у предварительно иммунизированных животных (через 0,5—2 часа), чем у неиммунизированных (через 4—24 часа). Параллельно изменяется и ультраструктура дендритных ретикулярных клеток. В опытах на кроликах показано, что в процессе формирования светлых центров дендритные ретикулярные клетки трансформируются из фибробластических ретикулярных клеток прилежащей лимфоцитарной короны. При этом ретикулярные клетки теряют активность щелочной фосфатазы (и, по-видимому, способность к волокнообразованию).
С возрастом в Ретикулярной ткани увеличивается количество волокнистых структур. Для костного мозга характерно необратимое замещение ретикулярной стромы жировой тканью и прекращение кроветворения. С возрастными и патологическими изменениями Р. т. тесно связано нарушение функции кроветворных и лимфоидных органов. При воздействии высоких доз ионизирующего излучения на организм Р. т. не регенерирует, а замещается фиброзной. Миелофиброз характеризуется интенсивной пролиферацией фиброгенных клеток и последующим массивным отложением коллагена I, III и IV типов. Показано участие ретикулярных клеток в синтезе фибриллярных белков амилоида (см. Амилоидоз). Поражение собственно ретикулярной стромы, сопровождающееся нарушением кроветворения, не всегда можно выявить морфологически. Поэтому важная роль отводится методам клонирования стромальных клеток in vitro, изучению их взаимодействия с кроветворными и лимфоидными клетками в жидкостных и агаровых культурах.
Таблица. Сравнительная характеристика активности ферментов в ретикулярных клетках и мононуклеарных фагоцитах (макрофагах) белой пульпы селезенки человека По данным Мюллер-Хермелинка (H. К. Muller-Hermelink) и др. (1974)
Активность ферментов в различных клетках лимфатического фолликула селезенки
Ретикулярные клетки лимфоцитарной короны
Дендритные ретикулярные клетки светлых центров
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом