
Рыхлая волокнистая соединительная ткань, межклеточное вещество
Межклеточное вещество, или внеклеточный матрикс (substantia intercellularis), соединительной ткани состоит из коллагеновых и эластических волокон, а также из основного (аморфного) вещества. Межклеточное вещество как у зародышей, так и у взрослых образуется, с одной стороны, путем секреции соединительнотканными клетками, а с другой — из плазмы крови, поступающей в межклеточные пространства.
В эмбриогенезе человека образование межклеточного вещества происходит начиная с 1—2-го месяца внутриутробного развития. В течение жизни межклеточное вещество постоянно обновляется — резорбируется и восстанавливается.
Коллагеновые волокна
Коллагеновые структуры, входящие в состав соединительных тканей организмов человека и животных, являются наиболее представительными ее компонентами, образующими сложную организационную иерархию. Основу всей группы коллагеновых структур составляет волокнистый белок — коллаген, который определяет свойства коллагеновых структр.
Коллаген составляет более 30% общей массы белков тела, причем около 40% его находится в коже, около 50% — в тканях скелета и 10% — в строме внутренних органов.
Коллагеновые волокна в составе разных видов соединительной ткани определяют их прочность. В рыхлой волокнистой соединительной ткани они располагаются в различных направлениях в виде волнообразно изогнутых, спиралевидно скрученных, округлых или уплощенных в сечении тяжей толщиной 1—3 мкм и более. Длина их различна.
Внутренняя структура коллагенового волокна определяется фибриллярным белком — коллагеном, который синтезируется на рибосомах гранулярной эндоплазматической сети фибробластов.
Различают более 20 типов коллагена, отличающихся молекулярной организацией, органной и тканевой принадлежностью. Например:
- коллаген I типа встречается главным образом в соединительной ткани кожи, сухожилиях, костях, роговице глаза, склере, стенке артерий и др.;
- коллаген II типа входит в состав гиалиновых и фиброзных хрящей, стекловидного тела и роговицы глаза;
- коллаген III типа находится в дерме кожи плода, в стенках крупных кровеносных сосудов, а также в ретикулярных волокнах (например, органов кроветворения);
- коллаген IV типа — встречается в базальных мембранах, капсуле хрусталика (в отличие от других типов коллагена он содержит гораздо больше боковых углеводных цепей, а также гидрооксилизина и гидрооксипролина);
- V тип коллагена присутствует в хорионе, амнионе, эндомизии, перимизии, коже, а также вокруг клеток (фибробластов, эндотелиальных, гладкомышечных), синтезирующих коллаген.
Коллаген IV и V типа не образует выраженных фибрилл.
В аминокислотном составе белка коллагена преобладает глицин (33% — каждая третья аминокислота), а также пролин и гидроксипролин.
Молекулы коллагена имеют длину около 280 нм и ширину 1,4 нм. Они построены из триплетов — трех полипептидных α-цепочек предшественника коллагена — проколлагена, свивающихся еще в клетке в единую тройную спираль. Проколлаген секретируется в межклеточное вещество. Проколлаген формирует первый, молекулярный, уровень организации коллагенового волокна.
Второй, надмолекулярный, уровень — внеклеточной организации коллагенового волокна — представляет агрегированные в длину и поперечно связанные с помощью водородных связей молекулы тропоколлагена, образующиеся путем отщепления концевых пептидов проколлагена. Сначала образуются протофибриллы, а 5—6 протофибрилл, скрепленных между собой боковыми связями, составляют микрофибриллы толщиной около 5 нм.
При участии гликозаминогликанов, также секретируемых фибробластами, формируется третий, фибриллярный, уровень организации коллагенового волокна. Коллагеновые фибриллы представляют собой поперечно исчерченные структуры толщиной в среднем 20—100 нм. Период повторяемости темных и светлых участков 64—67 нм. Каждая молекула коллагена в параллельных рядах, как полагают, смещена относительно соседней цепи на четверть длины, что служит причиной чередования темных и светлых полос. В темных полосах под электронным микроскопом видны вторичные тонкие поперечные линии, обусловленные расположением полярных аминокислот в молекулах коллагена.
Четвертый, волоконный, уровень организации — коллагеновое волокно, образующееся путем агрегации фибрилл, имеет толщину 1 — 10 мкм (в зависимости от топографии). В него входит различное количество фибрилл — от единичных до нескольких десятков. Волокна могут складываться в пучки (волокон) толщиной до 150 мкм.
Коллагеновые волокна отличаются малой растяжимостью и большой прочностью на разрыв. В воде толщина сухожилия в результате набухания увеличивается на 50%, а в разбавленных кислотах и щелочах — в 10 раз, но при этом волокно укорачивается на 30%. Способность к набуханию больше выражена у молодых волокон. При термической обработке в воде коллагеновые волокна образуют клейкое вещество (греч. kolla — клей), что и дало название этим волокнам.
Читайте также: Текстура вязаная ткань бесшовная текстура
Разновидностью коллагеновых волокон являются ретикулярные и преколлагеновые волокна. Последние представляют собой начальную форму образования коллагеновых волокон в эмбриогенезе и при регенерации. В их состав входят коллаген III типа и повышенное количество углеводов, которые синтезируются ретикулярными клетками органов кроветворения. Они образуют трехмерную сеть — ретикулум, что и обусловило их название.
Эластические волокна
Наличие эластических волокон в соединительной ткани определяет ее эластичность и растяжимость. По прочности эластические волокна уступают коллагеновым. Форма поперечного разреза волокон округлая и уплощенная. В рыхлой волокнистой соединительной ткани эластические волокна широко анастомозируют друг с другом. Толщина эластических волокон обычно меньше коллагеновых (0,2—1 мкм), но может достигать нескольких микрометров (например, в выйной связке). В составе эластических волокон различают микрофибриллярный и аморфный компоненты.
Основой эластических волокон является глобулярный гликопротеин — эластин, синтезируемый фибробластами и гладкими мышечными клетками. Для эластина характерно наличие двух производных аминокислот — десмозина и изодесмозина, которые участвуют в стабилизации молекулярной структуры эластина и придании ему способности к растяжению, эластичности.
Глобулярный белок эластин составляет первый, молекулярный, уровень организации эластического волокна.
Молекулы эластина вне клетки соединяются в цепочки — эластиновые протофибриллы — второй, надмолекулярный, уровень организации эластического волокна. Эластиновые протофибриллы в сочетании с гликопротеином (фибриллином) образуют микрофибриллы.
Четвертый уровень организации эластического волокна — волоконный. Зрелые эластические волокна содержат около 90 % аморфного компонента эластических белков (эластина) в центре, а по периферии — микрофибриллы.
Кроме зрелых эластических волокон, различают элауниновые и окситалановые волокна. В элауниновых волокнах соотношение микрофибрилл и аморфного компонента примерно равное, а окситалановые волокна состоят только из микрофибрилл.
Коллагеновые и эластические волокна в соединительной ткани образуют волокнистый остов с ориентированным, неориентированным и смешанным типами расположения волокон. Ориентированный (или оформленный) тип характеризуется параллельным расположением основной массы волокнистых структур (например, в сухожилиях, связках, фасциях). Неориентированный (или неоформленный) тип построен из волокон, не имеющих преимущественной ориентации (как например, дерма кожи). Смешанный тип волокнистого остова, как правило, имеет слоистое строение с чередованием направлений расположения волокнистых элементов.
Аморфный компонент межклеточного вещества
Клетки и волокна соединительной ткани заключены в аморфный компонент, или основное вещество (substantia fundamentalis). Эта гелеобразная субстанция представляет собой метаболическую, интегративно-буферную многокомпонентную среду, которая окружает клеточные и волокнистые структуры соединительной ткани, нервные и сосудистые элементы. В состав компонентов основного вещества входят белки плазмы крови, вода, неорганические ионы, продукты метаболизма паренхиматозных клеток, а также растворимые предшественники коллагена и эластина, протеогликаны, гликопротеины и комплексы, образованные ими. Все эти вещества находятся в постоянном движении и обновлении.
Гликозаминогликаны (ГАГ, ранее — «кислые мукополисахариды») — полисахаридные соединения, — линейные полимеры, построенные из повторяющихся дисахаридных единиц. Каждая из этих единиц содержит обычно гексуроновую кислоту и гексозамин (аминосахарид). Молекулы ГАГ содержат много гидроксильных, карбоксильных и сульфатных групп, имеющих отрицательный заряд, легко присоединяют молекулы воды и ионы, в частности Na+, и поэтому определяют гидрофильные свойства ткани. ГАГ проницаемы для кислорода и СО2, но предохраняют органы от проникновения чужеродных тел и белков. Гликозаминогликаны участвуют в формировании волокнистых структур соединительной ткани и их механических свойствах, репаративных процессах соединительной ткани, в регуляции роста и дифференцировке клеток. Среди гликозаминогликанов наиболее распространена в соединительной ткани гиалуроновая кислота, а также сульфатированные ГАГ: хондроитин-сульфаты (в хряще, коже, роговице), дерматансульфат (в коже, сухожилиях, в стенке кровеносных сосудов), кератансульфат, гепаринсульфат (в составе многих базальных мембран). Гепарин — гликозаминогликан, состоящий из глюкуроновой кислоты и гликозамина. В организме человека и животных он вырабатывается тучными клетками, является естественным противосвертывающим фактором крови.
Читайте также: В ателье было 15 метров ткани из нее сшили платье
Соединения белков с ГАГ носят название протеогликаны (ПГ). В соединительных тканях протеогликаны образуют сложные протеогликановые комплексы, определяющие во многом свойства всего межклеточного вещества.
В основе протеогликанового комплекса лежит длинная (около 1700 нм) линейная молекула гиалуроновой кислоты, к которой присоединяются 70-100 молекул протеогликанов.
Полианионная природа ПГ позволяет им обеспечивать транспорт воды, солей, аминокислот. Пространственная организация ПГ-комплексов образует своеобразное молекулярное сито, регулирующее диффузию воды и низкомолекулярных продуктов питания и обмена. Нарушение пористости этого «фильтра», например, при возрастном отношении гиалуроновой кислоты и хондроитинсульфатов в стенках сосудов является одной из предпосылок к развитию атеросклероза.
Гликопротеины (ГП, «неколлагеновые белки») — класс соединений белков с олигосахаридами (гексозаминами, гексозами, фукозами, сиаловыми кислотами). Гликопротеины входят в состав как волокон, так и аморфного вещества. К ним относятся:
• растворимые ГП, связанные с протеогликанами;
• ГП кальцинированных тканей;
• ГП, связанные с коллагеном (структурные ГП и ГП базальных мембран).
Гликопротеины играют большую роль в формировании структуры межклеточного вещества соединительной ткани и также определяют его функциональные особенности (примеры ГП: фибронектин, хондронектин, фибриллин, ламинин и др.).
Фибронектин — главный поверхностный гликопротеин фибробласта. В межклеточном пространстве он связан главным образом с интерстициальным коллагеном. Полагают, что фибронектин обусловливает липкость, подвижность, рост и специализацию клеток.
Фибриллин формирует микрофибриллы, усиливает связь между внеклеточными компонентами.
Ламинин — компонент базальной мембраны, состоящий из трех полипептидных цепочек, связанных: между собой дисульфидными соединениями, а также с коллагеном V типа и поверхностными рецепторами клеток.
Соединительная ткань состоит из коллагена эластина
• Основная функция эластина состоит в обеспечении эластических свойств тканей
• Мономеры эластина (известные как тропоэластиновые субъединицы) организованы в волокна, настолько прочные и устойчивые, что функционируют в течение всей жизни организмов
• Прочность этих волокон обусловлена образованием ковалентных сшивок между боковыми цепями лизина, находящегося в соседних мономерах эластина
• Эластичность волокон связана с наличием гидрофобных областей, которые при приложении силы растягиваются, а при снятии нагрузки спонтанно сокращаются
• Сборка волокон тропоэластина происходит во внеклеточном пространстве и находится под контролем трехступенчатого процесса
• Мутации в гене эластина являются причиной развития разнообразных патологических состояний, начиная от образования морщин на коже и заканчивая ранней детской смертностью
Как следует из названия, эластин представляет собой белок внеклеточного матрикса, главным образом, обеспечивающий эластические свойства тканей. Благодаря эластину, ткань может растягиваться и возвращаться к исходному размеру, дополнительно не расходуя энергию. Эластина особенного много в таких тканях, как кровеносные сосуды, кожа и легкие, где эластичность является критическим фактором, обеспечивающим правильное функционирование органа.
Например, эластичность кровеносных сосудов играет важную роль в поддержании надлежащего уровня кровяного давления, а эластичность легких с каждым вдохом обеспечивает их заполнение воздухом и последующее опорожнение.
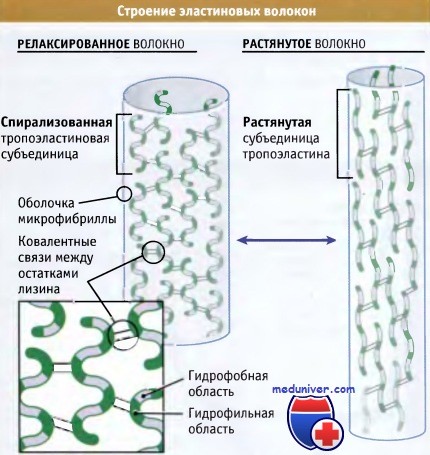
Схематическое строение релаксированных и растянутых эластиновых волокон.
Отметьте выраженные различия в структуре эластиновых субъединиц в каждом случае.
Детали строения субъединиц пока неизвестны.
Эластин синтезируется и секретируется фибробластами, представляющими собой один из наиболее распространенных типов клеток у животных, а также клетками гладкой мускулатуры. Эти клетки также секретируют коллагены, которые противостоят растяжению. В результате внеклеточный матрикс каждого органа характеризуется комбинацией свойств эластичности и прочности. Изменяя соотношение эластина и коллагена во внеклеточном матриксе, клетки могут регулировать эластические свойства органов.
Как следует из рисунка ниже, эластин организован в волокна, которые состоят из основной области эластиновых белков, окруженной оболочкой микрофибриллярных белков диаметром 10-12 нм. Основным компонентом этой оболочки является гликопротеин, связанный с микрофибриллами, который ассоциирован с мономером эластина и способствует его включению в более крупные эластиновые волокна. Эти волокна настолько прочны и стабильны, что функционируют на протяжении всей жизни организмов (т. е. не деградируя и не замещаясь). Присутствующий в этих волокнах эластин является наименее растворимым белком позвоночных.
Читайте также: Фольма ткань технические характеристики
Каким образом, эластин, обладая высокой прочностью и стабильностью, тем не менее, проявляет высокую эластичность? Ответ лежит в особенностях его структуры. Ген эластина содержит 36 экзонов, которые кодируют две совершенно разных последовательности аминокислот: некоторые из них проявляют гидрофильные свойства и содержат много остатков лизина, в то время, как другие обогащены гидрофобными аминокислотами, особенно глицином, пролином, аланином и валином.
Гидрофобные последовательности вкраплены среди гидрофильных областей, что обусловливает существование большого по размеру белка, обладающего двумя различными свойствами. Прочность эластиновых волокон, в основном определяется ковалентными связями, возникающими между боковыми цепями лизина в примыкающих друг к другу белках, подобно тому как это имеет место в коллагене. Наоборот, гидрофобные области обеспечивают эластичность за счет сворачивания молекулы в спираль в нерастянутом состоянии, и ее растяжения при приложении силы. При снятии напряжения эти области снова спонтанно сворачиваются. Даже после многолетних исследований биологи не пришли к заключению о точной конформации эластиновых белков в волокнах.
Сборка таких нерастворимых белков ставит перед клеткой особые проблемы. Если эти белки, до момента их секреции из клетки, спонтанно агрегируют, они могут повлиять на секрецию других белков, «закупоривая» секреторные пути, а также вызывать разрыв органелл или плазматической мембраны. Клетки синтезируют и секретируют эластиновые белки в виде мономеров, однако собирают волокна только во внеклеточном пространстве, после того как белки вышли из клетки, не нарушив ее внутреннюю среду.
Как иллюстрирует рисунок ниже, образование эластина включает три основных этапа:
• Вскоре после образования эластиновых мономеров (известных под названием тропоэластина) они связываются в ЭПР с белком-шапероном мол. массы 67 кДа. В течение всего процесса секреции этот шаперон остается связанным с тропоэластином и предотвращает агрегацию эластина в клетке.
• После завершения секреции комплекс с помощью шаперона удерживается на поверхности клетки до тех пор, пока он не вступит в контакт с оболочкой эластинового волокна. После этого вновь секрети-рованный тропоэластин встраивается в эластиновое волокно с участием сахарных компонентов его оболочки, которые замещают шапероны.
• Большая часть боковых цепей лизина, присутствующего в мономере тропоэластина, дезаминируется при действии фермента лизилоксидазы. При этом образуются аллизины, которые ковалентно связываются с аллизинами или немодифицированными лизинами, присутствующими в других эластиновых белках волокна. Название зрелый эластин используют для обозначения эластиновых белков, которые были модифицированы лизилоксидазой и образовали полимер.
Таким образом, трехэтапный метод обеспечивает полную сборку эластина только в тех местах клетки, где это является необходимым.
Как можно ожидать, изменения, наступающие в процессе сборки или функционировании эластина и эластиновых волокон, могут иметь серьезные последствия для клетки. Дряблость кожи, заболевание, которое выражается в утрате эластиновых волокон в коже и соединительной ткани, по тяжести проявления может варьировать от незначительного разрушения волокон и возникновения морщин до практически полной утраты волокон. У больных, в клетках которых мало эластиновых волокон или они вообще отсутствуют, не поддерживается целостность тканей, и они умирают в раннем детстве.
У больных с синдромом Вильямса образуются укороченные формы эластина, в которых отсутствуют некоторые домены связывания, и нарушен процесс организации в волокна. У этих больных развивается сильное сужение крупных артерий, по-видимому, обусловленное аномальным ростом вокруг артерий клеток гладкой мускулатуры. Это служит компенсаторной реакцией в ответ на утрату эластиновых волокон, обычно присутствующих в стенке артерий.
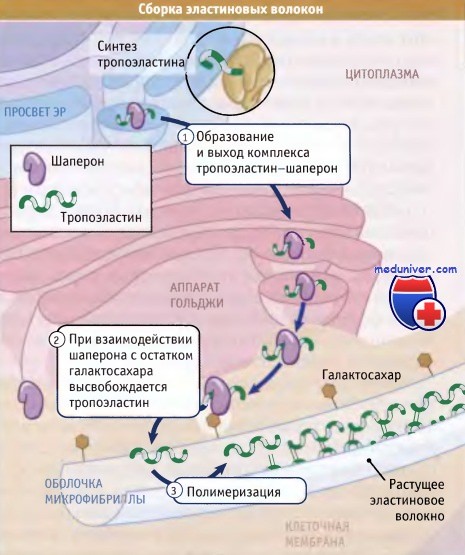
Во время транспорта к клеточной поверхности эластиновые мономеры (тропоэластины) связаны с шаперонами.
Связывание шаперонов с оболочкой микроволокна приводит к высвобождению мономеров эластина.
Полимеризация катализируется поперечными сшивками тропоэластинов, которые образуются при действии лизилоксидазы.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом