
Раздражители — живой клетки реагирует на действие раздр. изменения структур мембраны и протоионозотического свойством клетки. раздражимость — способность живой ткани отвечать раздражитель Раздражимость обладают нервная, мышечная и секреторная ткани их объединяют в понятиеВозбудимость.Возбуждение — это сложный биологический процесс, который характеризуется специфическим изменением процессов обмена веществ, теплообразования, временной деполяризацией мембраны клеток и проявляющийся специализированной реакцией ткани (сокращение мышцы, отделение секрета железой и т. д.).
физические(механические, температурные, звуковые, световые, электрические);
химические(щелочи, кислоты, гормоны, продукты обмена веществ и др.);
порог раздражения — минимальная сила раздражителя, которая способна вызвать возбуждение.
пороговые- миним.сила способная вызвать ответную реакцию
подпороговыми- если структура не отвечает на раздражение
более сильные — сверхпороговыми
По степени приспособленности биологических структур к их восприятию раздражители делятся на адекватные и неадекватные.
Первые попытки последовательной разработки учения о «животном электричестве» связаны с именем Л. Гальвани. Он обратил внимание на сокращение мышц препарата задних лапок лягушки, подвешенного на медном крючке, при прикосновении лапок к железным перилам балкона. На основании этих наблюдений Л. Гальвани пришел к выводу, что сокращение мышц лапок вызвано «животным электричеством», которое возникает в спинном мозге и передается по металлическим проводникам к мышцам лапки. Этот опыт в настоящее время известен как первый опыт Гальвани.
Функциональная активная структура — мембрана.
Функции – пограничная (цитоплазма от внеклеточной жидкости)
Биотрансформирующая (барьер для веществ идущих в клетку)
Рецепторная ( мембрана имеет набор структур участвующих в специальных определений физических и химических факторов)
Мембрана образует межклеточные контакты
Транспортная ( переносит разм в-ва в связи с изменение проницаемости)
В состав мембраны двойной слой фосфолипидов, в которой встроены белки и поверхность покрыта мукополисахаридами. Согласно жидкостно-мозаичной модели мембраны молекулы белка могут перемещаться в билипидном слое.
белки делятся на 5 классов:
насосы— расход метаб.энергии АТФ для перемещения ионов и молекул против концетракционных и электрохимических градиентов. Поддерживает необходимый концентрат ионов в клетке
каналы – сам. Транспортный механизм необходимый для переноса веществ через мембрану. Состоит из поры, воротного механизм, сенсора или индикатора и селективного фильтра. Пора это динамическое образование, которое может находиться в открытом или закрытом состоянии. Она образована белком с высокой каталитической активностью. Белок способен переноситься со скоростью в 200 раз больше чем скорость диффузии. Воротный механизм – находится на внутренней поверхности мембраны, состоит из отрицательно заряженных белков способные к конформации (изменение формы). Этот механизм изменяет величину потока ионов. Селективный фильтр на наружной поверхности мембраны , состоит из атомов О2, отрицательно заряженных притягив катионы, отталкиваются анионы. Формируется однонаправленное направление ионов. Сенсор – состоит из положительно заряденых белков.Классификация каналов: селективные – потенциалозависимыми., хемозависимые (рецепторуправляемые), по виду ионов: натриевые, калиевые, кальциевые, хлорные , по скорости: быстрые и медленные.
Неселективные (каналы утечки)
мембрана любой живой клетки проницаема для ионов оснований калия, кальция, натрия, и хлора, с помощью активных (идет против электрохимического градиента с затратой энергии и обязательным участием белков переносчиков) и пассивных ( диффузия , осмос, фильтрация; идентичен по концентрации градиенту без затраты энергии из большее в меньшее) транспортов
Состояние живой клетки: покой и возбуждение
Покой характеризуется Поляризация мембраны, т.е. наличие противоположных зарядов по обе стороны. В состоянии покоя мембрана в 10 раз более проницаема для ионов кальция, чем для ионов калия и хлора.
и ионы калия каналах по концентрационному градиенту выходит из клетки потому что в цитоплазме их в 50 раз больше, чем в внеклеточной жидкости. Это вызывает повышение электроотрицательные ионы расположения в цитоплазме. Белки цитоплазмы остаются на поверхности так как не могут пройти в цитоплазму. Клетка омывается внеклеточной жидкостью. Разница растворов ( цитоплазмы и внеклет) только в концентрации ионов. Растворы стремятся уровнять. Ионы калия выходят ионы натрия пытаются попасть каналы пропускают только мелкие ионы (белковые остатки, ионы на крупные) при помощи микроэлектор методики можно измерить величину заряда (мембр. Потенциал) за основу заряда внутренней поверхности мембранные это заряд называется мембранного потенциала покоя. Для разных тканей от 50 до 90 мВ ( со знаком «- «). Величина МПП : заряды, идущие через мембрану ионов и величиной потока ионов, направляющих поток ионов через ионный канал
Кроме того, концентрационного градиента имеется еще и сила, обеспечивающих перемещение ионов от одноименного к противоположному электрическому градиенту. А механизм, получил название натрий-калиевого насоса.Он обеспечивает активный транспорт ионов натрия во внеклеточную жидкость и введении в цитоплазму ионов калия. Переход из покоя в возбуждение характеризируется —деполяризациямембраны и выражается в виде генерации клеточного потенциала. В зависимости от силы раздражения потенциал может быть местным –локальный ответ. Такой потенциал не распространяется и затухает вблизи распространения. Возникаетдеполяризациисвязи с повышением проницаемости для ионов натрия идущих в клетку. Однако во время локального ответа мембранный потенциал не достигает критического уровня т.е. момента когда не распространяется возбуждение переходит в распространяющееся.
КУДне распространяющейся переход в распространяющееся . и возбуждение переходит на близ лежащие участки. Расстояние между мембранного потенциала и критического уровня называется пороговым популяциями. И возбуждение обратно существует . При действии на ткань раздражение пороговой силы повышает проницаемость для ионов натрия, т.к. открываются потенциально зависимые натрий- каналы и ионы натрия поступают внутрь клетки. Потенциально зависимые натрий- каналы имеют два типа ворот : М – активационными, и Н инактивационными
В состоянии покоя активация ворот закрыты, а инактивационные открыты. После нанесения раздражения активация ворот открыты и натрий устремляется внутрь клетки. К- каналы очень инертны
Постепенно натрий ток возрастает в 500 раз по сравнению состояния покоя и начинается преобладание на выход К-током. Процесс нарастает медленно до критического уровня деполяризации и процесс становится не управляем. Натрий лавиной устремляется в клетку и деполяризует мембрану зарядом «+». Ионный ток проходит «О» отметку, и заряжается «+». Натрий – каналы инактивируют (закрывают) и прекращают пропуск натрия, а калий только набирает и «+» заряду выход наружу. Достигает критического уровня К-каналы инактивируется, процесс замедляется. Насос выкачивает натрий и закачивает калий восстанавливает ионный баланс. Клетка вернулась в состояние покоя необходимо время для снижения силы натрий-калиевого насоса и натрий перекачивает на поверхность и возникает гиперполяризация и восстанавливается ионная ассиметрия.
Читайте также: Трикотаж вискоза ткань что это такое
Нанесение подпорогового раздражения возникает локальный ответ. Он способен к суммации (выход за критический уровень)
ВЫВОД: электрические явления на клетку мембраны . Для понятия реакции ткани на раздражение.
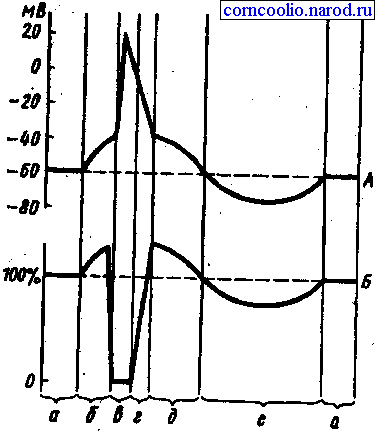
Рис. 2. Соотношение одиночного цикла возбуждения (А) и фаз возбудимости (Б). А: а — мембранный потенциал покоя; б — предспайк, локальный ответ, ВПСП; в — спайк, потенциал действия, деполяризация и инверсия; г — потенциал действия, реполяризация; д — отрицательный следовой потенциал, следовая деполяризация; е — положительный следовой потенциал, следовая гиперполяризация. Б: а — исходный уровень возбудимости; б — фаза первичной экзальтации, повышенная возбудимость; в — фаза абсолютной рефрактерности; г — фаза относительной рефрактернности; д — фаза вторичной экзальтации; е — фаза вторичной рефрактерности
Изменение возбудимости в разные фазы потенциального действия. В разные моменты токи движутся неравномерно. (отщепляясь АТФ выводит энергию и открывает ворота.
1. состояние локального ответа – ворота открыты мало АТФ не расщепилась, энергии очень много, наносим дополнительное раздражение – фаза возбудимости (супернормальная)
2 все каналы открыты, наносим дополнительное раздражение, ответа не будет (фаза абсолютной невозбудимости) фаза абсолютной рефрактернности
3 инактивация каналов локального ответа накапливается на мембране, наносит раздражение если раздражение пороговой силы ответа не будет если сверх пороговый то это фаза относительной рефрактернности
Отрицательно средовой потенциала. – знергия АТФ на мембрану накапливается следовательно на подпороговое раздражение получают максимальный ответ — фаза вторичной экзальтации. Гиперполяризация мембраны наносится раздражение продолжается выход ионов калия; заряд не изменяется ответ слабый (фаза субнормальной возбудимости
Физиология возбудимых тканей
1. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
1.1 Раздражимость и возбудимость живых систем
1.2 «Животное электричество». Опыты Гальвани и Матеучи
1.3 Мембранный потенциал покоя. Метод регистрации, механизмы происхождения и поддержания
1.4 Потенциал действия. Электрографические, электрохимические и функциональные проявления.
1.4.1. Электрографические проявления ПД
1.4.2. Электрохимические проявления ПД
1.4.4. Функциональные проявления ПД
1.5 Парабиоз. Оптимум и пессимум раздражения
2.1. Понятие и классификация нервных волокон
2.2 Свойства нервных волокон
2.3 Механизмы проведения возбуждения
3.1 Классификация синапсов
3.2 Этапы и механизмы синаптической передачи в химических синапсах
4.1 Виды и свойства рецепторов
4.2 Кодирование свойств раздражителей в рецепторах
4.3 Понятие о рецептивном поле и рефлексогенных зонах
1. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
1.1 Раздражимость и возбудимость живых систем
Биологические системы – организмы, органы, ткани и клетки – могут находиться в двух состояниях – покоя и активности.
Состояние покоя биосистемы можно наблюдать при отсутствии специальных раздражающих воздействий. Оно характеризуется относительным постоянством физиологических параметров и отсутствием проявлений специфических функций.
При изменениях внешней или внутренней среды (т.е. при воздействии раздражителей) биосистема может переходить в активное состояние. Способность всех живых систем реагировать на раздражители изменением своих свойств (обмен веществ и др.) называют раздражимостью. При этом раздражимость живых систем принципиально отличается от пассивной реактивности неживых тел (например, «реактивности» упругого мяча), поскольку энергия (сила) и форма ответных изменений не определяется энергией и формой воздействия.
Активное состояние некоторых тканей и клеток может сопровождаться не только изменением их свойств, но и проявлением специфической функции – реакцией. Реакция – изменение (усиление или ослабление) деятельности живой системы в ответ на раздражение. Реакции могут быть простые (генерация нервного импульса, сокращение, секреция) или сложные (пищедобывание).
Способность биосистемы отвечать на раздражение активной специфической реакцией называется возбудимостью. Клетки, способные к возбуждению называют возбудимыми. К ним относят нервные, мышечные, железистые клетки, а также элементы сенсорных рецепторов – нервные окончания и специальные рецепторные клетки.
Переход системы в активное состояние может быть как с положительным, так и с отрицательным знаком. В первом случае это увеличение уровня метаболизма, роста, повышение возбудимости по отношению к раздражителям. Это процессы возбуждения. Противоположные реакции – реакции торможения. Возбуждение и торможение представляют собой взаимопротивоположные и взаимосвязанные процессы.
Раздражение – процесс воздействия раздражителей на живой объект. Раздражители – это факторы внешней или внутренней среды, вызывающие переход биосистемы в активное состояние.
1. Физиология возбудимых тканей
В основе приспособительных реакций организма лежит раздражимость — способность реагировать на воздействия изменением структуры и функций. Раздражимостью обладают все клетки животных и растений. В ходе эволюции раздражимость некоторых тканей достигла наивысшего развития и трансформировалась в возбудимость (способность отвечать на раздражение возбуждением). К возбудимым относят нервную, мышечную и секреторную ткани. Возбудимость оценивают по порогу раздражения (минимальной силе раздражителя, которая способна вызвать возбуждение). Раздражители по их природе делят на физические, химические, биологические (вирусы, бактерии и др.), адекватные и неадекватные. Адекватными называют раздражители, к восприятию которых биологическая структура специально приспособлена. Поэтому пороговая сила адекватных раздражителей наименьшая. Например, адекватным для фоторецепторов является свет, для мышц — нервный импульс. Неадекватными называют раздражители, которые действуют на структуру, не приспособленную для их восприятия. Например, скелетная мышца реагирует сокращением и на электрические раздражения.
Биоэлектрические явления в возбудимых тканях. Возбуждение — это совокупность процессов, в результате которых кратковременная деполяризация цитоплазматической мембраны вызывает специализированную реакцию клетки (проведение нервного импульса, сокращение мышцы и т.д.).
Луиджи Гальвани обратил внимание на сокращение мышц препарата задних лапок, подвешенного на медном крючке, при соприкосновении с железными перилами балкона. На основании этого (первый опыт Гальвани) был сделан вывод, что сокращение вызвано «электричеством», которое передается по крючку и перилам от спинного мозга к мышцам. Однако физик А. Вольта предположил, что источником тока является не мозг, а потенциал в месте контакта разнородных металлов. В ответ на это Л. Гальвани стеклянным крючком набрасывал седалищный нерв на мышцы голени, что вызывало сокращение мышц (второй опыт или опыт без металлов) и доказывало существование «животного электричества». Позднее установлено, что клетки в покое внутри заряжены отрицательно по отношению к их поверхности. Этот потенциал покоя (ПП) составляет от 30 до 100 мВ.
Читайте также: Почему происходит отек тканей
В середине 20 века. А.Ходжкин, Э.Хаксли и Б.Катц создали мембранно-ионную теорию, согласно которой МП обусловлен разными концентрациями ионов калия, натрия и хлора по обе стороны клеточной мембраны. По сравнению с внеклеточной жидкостью, цитоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора. Высокая проницаемость мембраны для калия приводит к выходу части внутриклеточного калия в окружающую клетку среду и к появлению положительного заряда на наружной поверхности мембраны. Органические анионы, для которых мембрана непроницаема, создают на внутренней поверхности мембраны отрицательный заряд, а поддерживает высокие концентрации калия в клетке и натрия вне ее натрий-калиевый насос.
Раздражение возбудимой клетки может вызвать локальный ответ или потенциал действия. Локальный ответ возникает при подпороговых раздражениях. Он находится в прямой зависимости от силы стимула, локализуется на поверхности клетки только в месте ее раздражения и увеличивает возбудимость клетки. Потенциал действия (ПД) возникает под влиянием порогового или сверхпорогового раздражений. При этом проницаемость мембраны для натрия увеличивается и в результате проникновения натрия в клетку ее мембрана заряжается положительно по отношению к наружной среде. Затем закрываются натриевые и открываются дополнительные калиевые каналы. В результате выхода калия из клетки начинается восстанавление МП (реполяризация мембраны).
1. Предспайк (локальный ответ) — деполяризация мембраны до критического уровня.
2. Спайк — состоит из восходящей (деполяризация) и нисходящей (реполяризация) частей.
3. Следовой потенциал — состоит из следовой деполяризации и гиперполяризации.
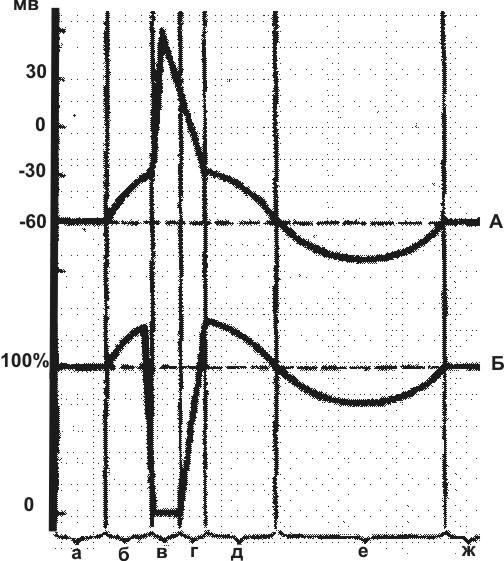
Рис. 1. Соотношение одиночного цикла возбуждения (А) и фаз возбудимости (Б).
А: а — потенциал покоя; б – предспайк (локальный ответ); в и г — спайк; д — следовая деполяризация; е — следовая гиперполяризация.
Б: а — исходный уровень возбудимости;
б — фаза повышенной возбудимости; в — фаза абсолютной рефрактерности; г — фаза относительной рефрактерности; д — фаза экзальтации; е — фаза субнормальной возбудимости.
Возбудимость в период предспайка повышена (фаза повышенной возбудимости) и даже слабый дополнительный стимул может вызвать формирование ПД. В период спайка мембрана не возбудима (абсолютная рефрактерность). Затем возбудимость постепенно восстанавливается (относительная рефрактерность). В это время для нового возбуждения нужно сверхпороговое раздражение. При следовой деполяризации возбудимость повышена (экзальтация), а при гиперполяризации — понижена (субнормальная возбудимость).
Законы раздражения отражают зависимость ответной реакции возбудимой ткани от силы раздражителя.
Закон «все или ничего»: подпороговые раздражители не вызывают ответной реакции («ничего»), а пороговые раздражители вызывают максимальный ответ («все»). По этому закону сокращается одиночное мышечное волокно и серце.
Закон силы: чем сильнее раздражение, тем больше ответная реакция. В соответствии с этим законом функционирует скелетная мышца. Она состоит из мышечных волокон с разной возбудимостью. На пороговые раздражители отвечают наиболее возбудимые волокна. Увеличение силы раздражителя дополнительно вовлекает в ответ волокна с меньшей возбудимостью и амплитуда сокращения мышцы растет.
Закон раздражения Дюбуа-Реймона: действие постоянного тока зависит от его силы и скорости нарастания. При медленном нарастании ткань приспосабливается к раздражителю (аккомодация) и возбуждение может не возникать.
Закон силы-времени отражает зависимость пороговой величины постоянного тока от времени его действия. Чем короче импульсы тока, тем меньшую раздражающую способность они имеют. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение которого ток, равный реобазе, вызывает возбуждение, называется полезным временем. Хронаксия — минимальное время, в течение которого ток, равный двум реобазам, вызывает реакцию.
Закон полярного действия постоянного тока: при замыкании тока возбуждение возникает под катодом, а при размыкании — под анодом.
Закон физиологического электротона: в области катода возбудимостьповышается (катэлектротон), а у анода — снижается (анэлектротон). При длительном действии постоянного тока возбудимость под катодом снижается (катодическая депрессия), а под анодом — растет (анодная экзальтация).
Нервное волокно обладает: возбудимостью, проводимостью и лабильностью. Возбуждения распространяется по нервному волокну только при его анатомической и физиологической целостности, не переходит на соседнее нервное волокно (закон изолированного проведения), не изменяется по амплитуде (закон незатухающего или бездекрементного проведения) и проводится в обе стороны от места раздражения (закон двустороннего проведения).
Возбужденный участок поверхности нервного волокна приобретает отрицательный заряд. Так как соседний невозбужденный участок заряжен положительно, то между ними потечет электрический ток. Это вызовет возбуждение покоящегося участка и тоже изменит его заряд. В конечном итоге возбуждение распространится по всей поверхности не покрытого миелиновой оболочкой (безмякотного) нервного волокна (рис. 2а.). В миелиновых (мякотных) отростках нейронов, возбуждение может возникать только в перехватах Ранвье. Поэтому оно распространяется скачками от одного перехвата к другому (рис. 2б.) и движется гораздо быстрее, чем в безмякотных волокнах.
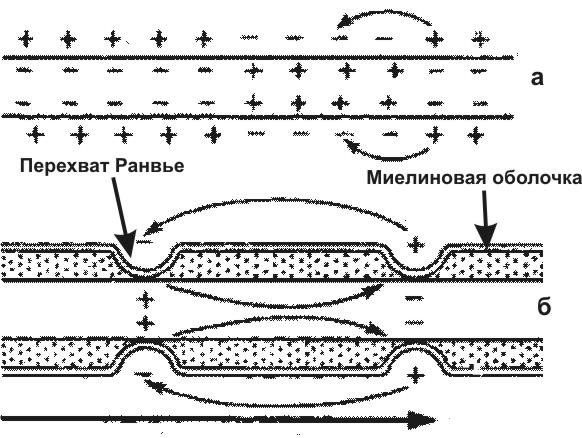
Рис. 2. Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б) нервных волокнах.
По диаметру и скорости проведения возбуждения нервные волокна делят на типы А, В и С. Самые толстые волокна типа А (диаметр 12-22 мкм) с наибольшей скоростью (70-120 м/с) проводят возбуждение от мозга к скелетным мышцам и от рецепторов мышц к мозгу. От многих других рецепторов идут волокна типа А с несколько меньшими диаметром (от 8 до 1 мкм) и скоростью проведения возбуждения (5-70 м/с). К волокнам типа В относятся преганглионарные вегетативные волокна (диаметр — 1-3,5 мкм, скорость проведения возбуждения — 3-18 м/с). Только волокна типа С являются безмякотными (их диаметр 0,5-2 мкм, скорость проведения возбуждения менее 3 м/с). Они являются постганглионарными симпатическими волокнами, а также идут от болевых рецепторов, части терморецепторов и рецепторов давления.
Нервные волокна обладают лабильностью (функциональной подвижностью). Ее измеряют по максимальному количеству возбуждений, которое способно воспроизвести нервное волокно. У нервных волокон лабильность выше (до 1000 Гц) чем в других возбудимых структурах. Если нерв повредить (химическим веществом, нагреванием, охлаждением или током) не нарушая анатомическую целостность, то в нем развивается состояние парабиоза. При этом последовательно сменяются уравнительная, парадоксальная и тормозная фазы. В уравнительную фазу — при редких раздражениях все импульсы проводятся через поврежденный участок, а при высоком ритме только часть. В парадоксальную — ответная реакция на частые раздражения меньше, чем на редкие. В тормозную — нерв не проводит любые возбуждения. При выходе из парабиоза наблюдаются те же фазы, но в обратной последовательности.
Читайте также: Покровная ткань растений выполняет функции строение
Все мышцы обладают возбудимостью (способностью возбуждаться при действии раздражителей), проводимостью (способностью проводить возбуждение) и сократимостью (способностью изменять свою длину или напряжение при возбуждении). Для сердечных и части гладких мышечных волокон, дополнительно к перечисленным свойствам, характерна автоматия (способность к самопроизвольному возбуждению). Уникальным свойством всех гладких мышц является пластичность (способность долго сохранять приданную им длину).
Сила мышцы определяется максимальным грузом, который она может поднять, а работа — произведением величины поднятого груза на высоту подъема. Максимальная работа производится при средних величинах нагрузок. При изотоническом сокращении мышцы изменяется ее длина, а напряжение постоянно (так сокращаются мышцы при отсутствии сопротивления изменению длины). При изометрическом сокращении длина мышцы постоянна, а ее напряжение растет (например, при попытке поднять чрезмерный груз). В естественных условиях наблюдаются смешанные сокращения (изменяются и длина и напряжение мышцы).
Один стимул вызывает одиночное мышечное сокращение. В нем выделяют: латентный период (время от начала действия раздражителя до начала ответной реакции), фазу укорочения и фазу расслабления. Если каждый последующий стимул поступает к скелетной мышце в период ее укорочения — возникает гладкий тетанус, а в фазу расслабления — зубчатый тетанус. В естественных условиях к скелетной мышце поступает такая серия импульсов, на которую мышца отвечает гладким тетанусом. Его амплитуда, как правило, выше амплитуды одиночного сокращения. Н.Е. Введенский объяснил это оптимумом и пессимумом. Оптимум — частота, при которой раздражения наносятся в фазу повышенной возбудимости (тетанус максимальный). Пессимум — частота, при которой новое раздражение наносится в фазу пониженной возбудимости (тетанус – минимален).
При изучении скелетных и сердечных мышц в поляризованном свете, видны чередующиеся зоны с различной оптической плотностью (рис. 3). Это позволило разделить сократительные элементы поперечнополосатых мышечных волокон (миофибрилл) на функциональные единицы — саркомеры (участки между соседними Z-мембранами).
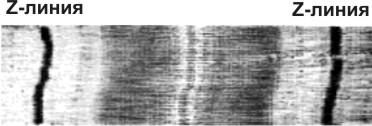
Рис. 3. Микрофотография участка поперечнополосатой мышцы.
Характерная оптическая плотность участков саркомера обусловлена особенностями расположения в них (рис. 4) сократительных белков (актина и миозина).

Рис. 4. Схема расположения сократительных белков в саркомере (в Н-зоне миозиновые фибриллы не имеют мостиков).
Около Z-мембран содержится актин (светлая область – изотропный или I-диск). Ближе к середине саркомера видны темные (анизотропные) А-диски с миозином и актином. В центре саркомера проходит М-линия. По обе стороны от нее расположен миозин (просветление или Н-зона). Актин с одной стороны прикреплен к Z-мембране, а с другой свободен и заканчивается между миозиновыми волокнами в области А-диска (на границе с Н-зоной). Оба конца миозиновой нити свободны.
При сокращении уменьшается ширина только изотропных дисков. Миозиновые нити при этом могут достигать своими концами Z-пластинок, а длина саркомеров — укорачиваться на 30-50%.
Механизм мышечного сокращения. Практически на всей миозиновой нити имеются боковые мостики (отсутствуют только около М-линии). После сцепления с актином они изменяют угол наклона (используя энергию АТФ), что продвигает сократительные белки относительно друг друга (саркомер укорачивается). Затем актин соединяется с другим мостиком миозина и совершается дальнейшее продвижение.
В покое соединению актина с миозином мешают белки тропонин и тропомиозин. При возбуждении они «нейтрализуются» поступающим в саркоплазму (цитоплазму мышечной клетки) кальцием и начинается взаимодействие сократительных белков. Прекращение возбуждения приводит к удалению кальция из саркоплазмы, тропонин с тропомиозином разрушают комплекс актина и миозина – мышца расслабляется.
В скелетных мышцах используется кальций из внутриклеточных цистерн саркоплазматической сети. Гладкие мышцы получают кальций только из межклеточных пространств, а сердечная мускулатура использует оба источника данного иона. Использование только внутриклеточных источников кальция позволяет скелетным мышцам сокращаться и расслабляться с наибольшей скоростью, а гладкая мускулатура — медленно изменяет свой тонус.
Синапс — функциональное соединение между нейроном и другими клетками. Существуют электрические и химические синапсы. Электрическим синапсам свойственно низкое электрическое сопротивление в области контакта клеток и ПД быстро передается на соседнюю мембрану. Химические синапсы состоят из пресинаптической мембраны, постсинаптической мембраны и синаптической щели (рис. 5.). Под влиянием нервных импульсов, в химических синапсах происходит высвобождение химического вещества — медиатора (например, ацетилхолина, норадреналина) из пресинаптических утолщений аксона в синаптическую щель и его взаимодействие с рецепторами на постсинаптической мембране. В возбуждающих синапсах это приводит к возникновению возбуждающего (ВПСП), а в тормозных — тормозного (ТПСП) постсинаптических потенциалов. После того как медиатор выполнил свою функцию, он разрушается или поглощается клетками.
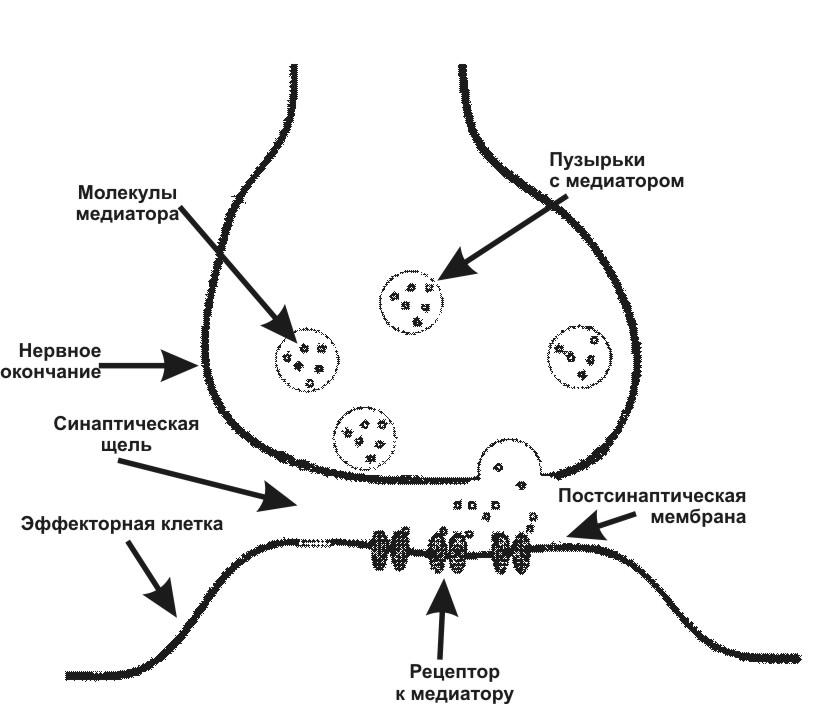
Рис. 5. Схема строения химического синапса.
За пределами ЦНС наиболее распространен медиатор ацетилхолин. Он облегчает проведение возбуждения через вегетативные ганглии, повышает секрецию адреналина надпочечниками и соляной кислоты желудочными железами, угнетает работу сердца, вызывает сокращение гладких мышц некоторых внутренних органов и экзокринных желез. В гладких мышцах бронхов, кишечника, мочевого пузыря, матки, круговой и цилиарной мышцах глаза ацетилхолин приводит, соответственно, к бронхоспазму, усилению перистальтики кишечника и желудка (при расслаблении сфинктеров), сокращению мочевого пузыря и сужению зрачка.
По расположению синапсы делят на периферические (нервно-мышечные, рецепторно-нейрональные и т.д.); центральные (аксосоматические, аксодендритные и др.); по знаку действия (возбуждающие и тормозящие) и по выделяемому медиатору (холинергические, адренергические и др.).
Химические синапсы проводят возбуждение в одном направлении; передают возбуждение медленнее, чем по нервному волокну (синаптическая задержка); имеют низкую лабильность, а также высокую утомляемость и чувствительность к химическим веществам.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом