
В возбудимой ткани невозможно вызвать новый потенциал действия, пока мембрана возбужденного волокна остается деполяризованной в связи с развитием предыдущего потенциала действия. Это связано с тем, что вскоре после возникновения потенциала действия натриевые каналы (или кальциевые каналы, или оба типа каналов) инактивируются, и возбуждающий сигнал любой силы, действующий на ткань в этот момент, не может открыть инактивационные ворота. Единственным условием для их повторного открытия является возврат мембранного потенциала к исходному или близкому к исходному уровню. Затем в течение небольшой доли секунды инактивационные ворота каналов открываются, и становится возможным развитие нового потенциала действия.
Период, в течение которого нельзя вызвать второй потенциал действия даже сильным стимулом, называют абсолютным рефрактерным периодом. Для крупных миелинизированных нервных волокон этот период составляет примерно 1/2500 сек. Легко рассчитать, что такое волокно может максимально передавать около 2500 имп/сек.
Кроме факторов, повышающих возбудимость нервного волокна, существуют так называемые мембраностабилизирующие факторы, способные снижать возбудимость. Например, высокая концентрация ионов кальция во внеклеточной жидкости уменьшает проницаемость мембраны для ионов натрия, снижая возбудимость. В связи с этим ионы кальция называют стабилизатором.
Местные анестетики. К наиболее важным стабилизаторам относят многие вещества, используемые в клинике в качестве местных анестетиков, в состав которых входят прокаин и тетракаин. Большинство из них действуют непосредственно на активационные ворота натриевых каналов, затрудняя их открытие, что сопровождается снижением возбудимости мембран. После снижения возбудимости до уровня, при котором отношение амплитуды потенциала действия к порогу возбуждения (называемое фактором надежности) опускается ниже 1,0, импульсы по анестезированному нерву не проходят.
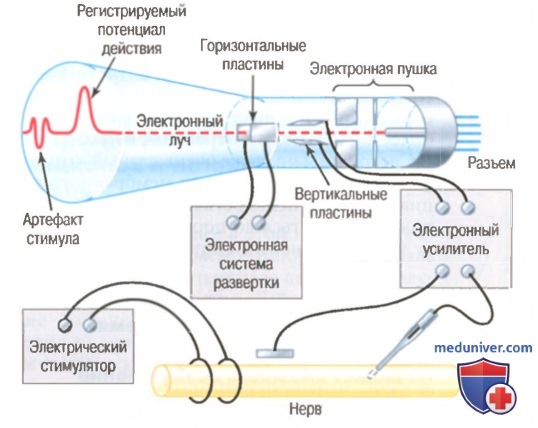
Катодно-лучевой осциллоскоп для регистрации быстрых потенциалов действия.
Катодный осциллограф. Ранее в этой главе мы отмечали, что изменения мембранного потенциала во время генерации потенциала действия происходят чрезвычайно быстро. Действительно, развитие большей части комплекса потенциала действия в крупных нервных волокнах занимает менее 1/1000 сек. На некоторых рисунках данной главы показан электроизмерительный прибор, регистрирующий эти изменения потенциалов. Однако ясно, что реакции любого прибора, способного регистрировать потенциалы действия, должны быть чрезвычайно быстрыми. Для практических целей единственным широко используемым прибором, способным точно реагировать на быстрые изменения мембранного потенциала, является катодно-лучевой осциллоскоп.
На рисунке показаны основные компоненты катодно-лучевого осциллоскопа. Катодно-лучевая трубка состоит из электронной пушки и флюоресцентного экрана, который «бомбардируется» электронами. При ударе электронов о поверхность экрана флюоресцентный материал светится. Если луч электронов перемещается по экрану, вместе с ним движется яркое пятно света, оставляя на экране флюоресцирующую линию.
Кроме электронной пушки и флюоресцентной поверхности, катодно-лучевая трубка снабжена двумя парами электрически заряженных пластин. Одна пара расположена с обеих сторон от электронного луча, а другая — сверху и снизу. Соответствующие электронные усилители изменяют напряжение на этих пластинах таким образом, что электронный луч отклоняется вверх или вниз в ответ на электрические сигналы, приходящие от регистрирующих электродов. Под влиянием внутреннего электронного блока осциллоскопа луч электронов перемещается по экрану горизонтально с постоянной скоростью. При этом регистрируется кривая, которую видно на экране катодно-лучевой трубки рисунка, с временной разверткой по горизонтали и изменениями потенциалов, регистрируемых отводящими электродами, по вертикали.
На левом конце кривой виден небольшой артефакт стимула, связанный с электрическим стимулом, который используется для вызова потенциала действия, справа на кривой — сам потенциал действия.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Фазовые изменения возбудимости при потенциале действия
В процессе развития потенциала действия происходит изменение возбудимости возбудимых тканей. Эти изменения носят фазовый характер, связаны с изменениями заряда мембраны и изменениями ее проводимости для ионов (рис. 15).

В период медленной деполяризации (1) начинают открываться натриевые каналы и возбудимость возрастает, что приводит к снижению порога возбуждения. ( Порог возбуждения характеризуется величиной деполяризации от Ео до Ек ). Для открытия дополнительного количества натриевых каналов необходима меньшая сила внешнего раздражителя. Эта фаза повышенной возбудимости (I).
В период быстрой деполяризации (2) все натриевые каналы от- крыты и активированы, входящий ток натрия максимален. Любое внешнее раздражение не создает дополнительной деполяризации. Эта фаза носит название фазы абсолютной рефрактерности (невозбудимости) (II).
В период реполяризации (3) часть натриевых каналов инактивирована медленной h частицей, но часть после инактивации уже перешла в закрытое активированное состояние. Т.е. внешнее воздействие способно перевести их в открытое активированное состояние, а значит вызвать дополнительную деполяризацию мембраны. Сила внешнего раздражения должна быть значительно больше пороговой. Эта фаза носит название относительной рефрактерности (III).
В период следовой деполяризации или следового отрицательного потенциала (4) в связи с остаточно увеличенной натриевой проводимостью мембраны уменьшается порог возбуждения, а возбудимость воз- растает. Эта фаза носит название фазы супернормальной возбудимости или фазы экзальтации (IV). В эту фазу возбуждение может быть вы- звано действием подпорогового стимула.
В период следовой гиперполяризации или следового положительного потенциала (5), обусловленной повышенным током калия, порог возбуждения увеличивается, а возбудимость уменьшается. Эта фаза субнормальной возбудимости (V). Возбуждение может быть вызвано только сверхпороговыми раздражителями.
Фазы возбудимости при возбуждении.
При развитии потенциала действия происходит изменение возбудимости ткани, причем, это изменение протекает по фазам (рис.2).

Рис. 2. Соотношение одиночного цикла возбуждения (А) и фаз
А: а — мембранный потенциал покоя; б — предспайк, локальный ответ, ВПСП; в —
спайк, потенциал действия, деполяризация и инверсия; г — потенциал действия, реполяризации; д — отрицательный следовой потенциал, следовая деполяризация; е – положительный следовой потенциал, следовая гиперполяризация. Б: а — исходный уровень возбудимости; б — фаза первичной экзальтации, повышенная возбудимость; в — фаза абсолютной рефрактерности; г — фаза относительной рефрактерности; д — фаза вторичной . ; е — фаза вторичной рефрактерности.
Читайте также: Чем отличается пвх от прорезиненной ткани
Состоянию исходной поляризации мембраны, которую отражает мембранный потенциал покоя, соответствует исходное состояние ее возбудимости и, следовательно, клетки — это нормальный уровень возбудимости. В период локального ответа возбудимость ткани повышена, эта фаза возбудимости получила название первичной экзальтации. Во время развития локального ответа мембранный потенциал покоя приближается к критическому уровню деполяризации и для достижения последнего достаточна сила раздражителя меньшая, чем пороговая (подпороговая).
В период развития пикового потенциала идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны и она утрачивает способность отвечать возбуждением на раздражители даже сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности (абсолютной невозбудимости). Ома длится до конца перезарядки мембраны. Абсолютная рефрактерность, т. е. полная не возбудимость мембраны возникает в связи с тем, что натриевые каналы в начале полностью открываются, а затем инактивируются.
После окончания фазы перезарядки мембраны возбудимость ее постепенно восстанавливается до исходного уровня — фаза относительной рефрактерности. Она продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Так как и этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена, и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых.
Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости — фаза вторичной экзальтации. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, но сравнению с состоянием покоя (исходной поляризацией), то порог раздражения снижен, т. е. возбудимость повышена. В эту фазу новое возбуждение может возникнуть при действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы неполностью. В Период развития положительного следового потенциала возбудимость ткачи понижена — фаза вторичной рефрактерности. В эту фазу мембранный Потенциал увеличивается (состояние гиперполяризации мембраны), удаляясь от критического уровня деполяризации, порог раздражения повышается и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины. Гиперполяризация мембраны развивается вследствие трех причин: во-первых, продолжающимся выходом ионов калия; во-вторых, открытием, возможно, каналов для хлора и поступление этиx ионов в цитоплазму клетки; в-третьих, усиленной работой натрий-калиевого насоса.
Закон силы-длительности: раздражающее действие постоянного тока зависит не только от его величины, но и от времени, в течение которого он Действует. Чем больше ток, тем меньше времени он должен действовать для возникновения возбуждения.
Исследования зависимости силы-длительности показали, что последняя имеет гиперболический характер (рис. 3).

Рис3. Графическое выражение законы силы-длительности.
Из этого следует, что сок ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он не действовал, и чем короче импульсы тока, тем меньшую раздражающую способность они имеют. Причиной такой’ зависимости является мембранная емкость. Очень «короткие» токи просто не успевают разрядить эту емкость до критического уровня деполяризации. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение которого действует ток, равный реобазе, и вызывает возбуждение, называется полезным временем.
В связи с тем, что определение этого времени затруднено, было введено понятие хронаксия — минимальное время, в течение которого ток, равный двум реобазам, должен действовать на ткань, чтобы вызвать ответную реакцию. Определение хронаксии — хронаксиметрия — находит применение в клинике. Электрический ток, приложенный к мышце, проходит через как мышечные, так и нервные волокна, и их окончания, находящиеся в этой мышце. Так как хронаксия нервных волокон значительно меньше хронаксии мышечных волокон, то при исследовании хронаксии мышцы практически получают хронаксию нервных волокон. Если нерв поврежден или произошла гибель соответствующих мотонейронов спинного мозга (это имеет место при полимиелите и некоторых других заболеваниях), то происходит перерождение нервных волокон и тогда определяется хронаксия уже мышечных волокон, которая имеет большую величину, чем нервных волокон.
1. Физиология возбудимых тканей
В основе приспособительных реакций организма лежит раздражимость — способность реагировать на воздействия изменением структуры и функций. Раздражимостью обладают все клетки животных и растений. В ходе эволюции раздражимость некоторых тканей достигла наивысшего развития и трансформировалась в возбудимость (способность отвечать на раздражение возбуждением). К возбудимым относят нервную, мышечную и секреторную ткани. Возбудимость оценивают по порогу раздражения (минимальной силе раздражителя, которая способна вызвать возбуждение). Раздражители по их природе делят на физические, химические, биологические (вирусы, бактерии и др.), адекватные и неадекватные. Адекватными называют раздражители, к восприятию которых биологическая структура специально приспособлена. Поэтому пороговая сила адекватных раздражителей наименьшая. Например, адекватным для фоторецепторов является свет, для мышц — нервный импульс. Неадекватными называют раздражители, которые действуют на структуру, не приспособленную для их восприятия. Например, скелетная мышца реагирует сокращением и на электрические раздражения.
Биоэлектрические явления в возбудимых тканях. Возбуждение — это совокупность процессов, в результате которых кратковременная деполяризация цитоплазматической мембраны вызывает специализированную реакцию клетки (проведение нервного импульса, сокращение мышцы и т.д.).
Луиджи Гальвани обратил внимание на сокращение мышц препарата задних лапок, подвешенного на медном крючке, при соприкосновении с железными перилами балкона. На основании этого (первый опыт Гальвани) был сделан вывод, что сокращение вызвано «электричеством», которое передается по крючку и перилам от спинного мозга к мышцам. Однако физик А. Вольта предположил, что источником тока является не мозг, а потенциал в месте контакта разнородных металлов. В ответ на это Л. Гальвани стеклянным крючком набрасывал седалищный нерв на мышцы голени, что вызывало сокращение мышц (второй опыт или опыт без металлов) и доказывало существование «животного электричества». Позднее установлено, что клетки в покое внутри заряжены отрицательно по отношению к их поверхности. Этот потенциал покоя (ПП) составляет от 30 до 100 мВ.
В середине 20 века. А.Ходжкин, Э.Хаксли и Б.Катц создали мембранно-ионную теорию, согласно которой МП обусловлен разными концентрациями ионов калия, натрия и хлора по обе стороны клеточной мембраны. По сравнению с внеклеточной жидкостью, цитоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора. Высокая проницаемость мембраны для калия приводит к выходу части внутриклеточного калия в окружающую клетку среду и к появлению положительного заряда на наружной поверхности мембраны. Органические анионы, для которых мембрана непроницаема, создают на внутренней поверхности мембраны отрицательный заряд, а поддерживает высокие концентрации калия в клетке и натрия вне ее натрий-калиевый насос.
Читайте также: Какую ткань выбрать для платья с запахом
Раздражение возбудимой клетки может вызвать локальный ответ или потенциал действия. Локальный ответ возникает при подпороговых раздражениях. Он находится в прямой зависимости от силы стимула, локализуется на поверхности клетки только в месте ее раздражения и увеличивает возбудимость клетки. Потенциал действия (ПД) возникает под влиянием порогового или сверхпорогового раздражений. При этом проницаемость мембраны для натрия увеличивается и в результате проникновения натрия в клетку ее мембрана заряжается положительно по отношению к наружной среде. Затем закрываются натриевые и открываются дополнительные калиевые каналы. В результате выхода калия из клетки начинается восстанавление МП (реполяризация мембраны).
1. Предспайк (локальный ответ) — деполяризация мембраны до критического уровня.
2. Спайк — состоит из восходящей (деполяризация) и нисходящей (реполяризация) частей.
3. Следовой потенциал — состоит из следовой деполяризации и гиперполяризации.
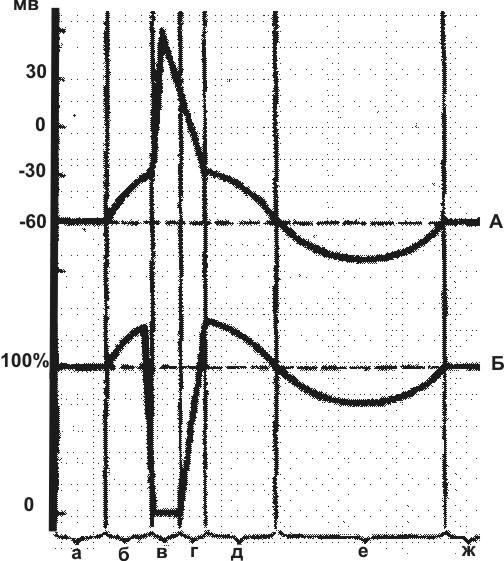
Рис. 1. Соотношение одиночного цикла возбуждения (А) и фаз возбудимости (Б).
А: а — потенциал покоя; б – предспайк (локальный ответ); в и г — спайк; д — следовая деполяризация; е — следовая гиперполяризация.
Б: а — исходный уровень возбудимости;
б — фаза повышенной возбудимости; в — фаза абсолютной рефрактерности; г — фаза относительной рефрактерности; д — фаза экзальтации; е — фаза субнормальной возбудимости.
Возбудимость в период предспайка повышена (фаза повышенной возбудимости) и даже слабый дополнительный стимул может вызвать формирование ПД. В период спайка мембрана не возбудима (абсолютная рефрактерность). Затем возбудимость постепенно восстанавливается (относительная рефрактерность). В это время для нового возбуждения нужно сверхпороговое раздражение. При следовой деполяризации возбудимость повышена (экзальтация), а при гиперполяризации — понижена (субнормальная возбудимость).
Законы раздражения отражают зависимость ответной реакции возбудимой ткани от силы раздражителя.
Закон «все или ничего»: подпороговые раздражители не вызывают ответной реакции («ничего»), а пороговые раздражители вызывают максимальный ответ («все»). По этому закону сокращается одиночное мышечное волокно и серце.
Закон силы: чем сильнее раздражение, тем больше ответная реакция. В соответствии с этим законом функционирует скелетная мышца. Она состоит из мышечных волокон с разной возбудимостью. На пороговые раздражители отвечают наиболее возбудимые волокна. Увеличение силы раздражителя дополнительно вовлекает в ответ волокна с меньшей возбудимостью и амплитуда сокращения мышцы растет.
Закон раздражения Дюбуа-Реймона: действие постоянного тока зависит от его силы и скорости нарастания. При медленном нарастании ткань приспосабливается к раздражителю (аккомодация) и возбуждение может не возникать.
Закон силы-времени отражает зависимость пороговой величины постоянного тока от времени его действия. Чем короче импульсы тока, тем меньшую раздражающую способность они имеют. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение которого ток, равный реобазе, вызывает возбуждение, называется полезным временем. Хронаксия — минимальное время, в течение которого ток, равный двум реобазам, вызывает реакцию.
Закон полярного действия постоянного тока: при замыкании тока возбуждение возникает под катодом, а при размыкании — под анодом.
Закон физиологического электротона: в области катода возбудимостьповышается (катэлектротон), а у анода — снижается (анэлектротон). При длительном действии постоянного тока возбудимость под катодом снижается (катодическая депрессия), а под анодом — растет (анодная экзальтация).
Нервное волокно обладает: возбудимостью, проводимостью и лабильностью. Возбуждения распространяется по нервному волокну только при его анатомической и физиологической целостности, не переходит на соседнее нервное волокно (закон изолированного проведения), не изменяется по амплитуде (закон незатухающего или бездекрементного проведения) и проводится в обе стороны от места раздражения (закон двустороннего проведения).
Возбужденный участок поверхности нервного волокна приобретает отрицательный заряд. Так как соседний невозбужденный участок заряжен положительно, то между ними потечет электрический ток. Это вызовет возбуждение покоящегося участка и тоже изменит его заряд. В конечном итоге возбуждение распространится по всей поверхности не покрытого миелиновой оболочкой (безмякотного) нервного волокна (рис. 2а.). В миелиновых (мякотных) отростках нейронов, возбуждение может возникать только в перехватах Ранвье. Поэтому оно распространяется скачками от одного перехвата к другому (рис. 2б.) и движется гораздо быстрее, чем в безмякотных волокнах.
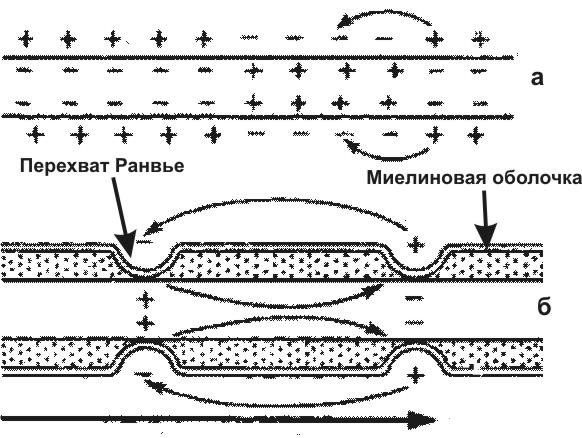
Рис. 2. Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б) нервных волокнах.
По диаметру и скорости проведения возбуждения нервные волокна делят на типы А, В и С. Самые толстые волокна типа А (диаметр 12-22 мкм) с наибольшей скоростью (70-120 м/с) проводят возбуждение от мозга к скелетным мышцам и от рецепторов мышц к мозгу. От многих других рецепторов идут волокна типа А с несколько меньшими диаметром (от 8 до 1 мкм) и скоростью проведения возбуждения (5-70 м/с). К волокнам типа В относятся преганглионарные вегетативные волокна (диаметр — 1-3,5 мкм, скорость проведения возбуждения — 3-18 м/с). Только волокна типа С являются безмякотными (их диаметр 0,5-2 мкм, скорость проведения возбуждения менее 3 м/с). Они являются постганглионарными симпатическими волокнами, а также идут от болевых рецепторов, части терморецепторов и рецепторов давления.
Нервные волокна обладают лабильностью (функциональной подвижностью). Ее измеряют по максимальному количеству возбуждений, которое способно воспроизвести нервное волокно. У нервных волокон лабильность выше (до 1000 Гц) чем в других возбудимых структурах. Если нерв повредить (химическим веществом, нагреванием, охлаждением или током) не нарушая анатомическую целостность, то в нем развивается состояние парабиоза. При этом последовательно сменяются уравнительная, парадоксальная и тормозная фазы. В уравнительную фазу — при редких раздражениях все импульсы проводятся через поврежденный участок, а при высоком ритме только часть. В парадоксальную — ответная реакция на частые раздражения меньше, чем на редкие. В тормозную — нерв не проводит любые возбуждения. При выходе из парабиоза наблюдаются те же фазы, но в обратной последовательности.
Читайте также: Костюмы из пикачу ткани
Все мышцы обладают возбудимостью (способностью возбуждаться при действии раздражителей), проводимостью (способностью проводить возбуждение) и сократимостью (способностью изменять свою длину или напряжение при возбуждении). Для сердечных и части гладких мышечных волокон, дополнительно к перечисленным свойствам, характерна автоматия (способность к самопроизвольному возбуждению). Уникальным свойством всех гладких мышц является пластичность (способность долго сохранять приданную им длину).
Сила мышцы определяется максимальным грузом, который она может поднять, а работа — произведением величины поднятого груза на высоту подъема. Максимальная работа производится при средних величинах нагрузок. При изотоническом сокращении мышцы изменяется ее длина, а напряжение постоянно (так сокращаются мышцы при отсутствии сопротивления изменению длины). При изометрическом сокращении длина мышцы постоянна, а ее напряжение растет (например, при попытке поднять чрезмерный груз). В естественных условиях наблюдаются смешанные сокращения (изменяются и длина и напряжение мышцы).
Один стимул вызывает одиночное мышечное сокращение. В нем выделяют: латентный период (время от начала действия раздражителя до начала ответной реакции), фазу укорочения и фазу расслабления. Если каждый последующий стимул поступает к скелетной мышце в период ее укорочения — возникает гладкий тетанус, а в фазу расслабления — зубчатый тетанус. В естественных условиях к скелетной мышце поступает такая серия импульсов, на которую мышца отвечает гладким тетанусом. Его амплитуда, как правило, выше амплитуды одиночного сокращения. Н.Е. Введенский объяснил это оптимумом и пессимумом. Оптимум — частота, при которой раздражения наносятся в фазу повышенной возбудимости (тетанус максимальный). Пессимум — частота, при которой новое раздражение наносится в фазу пониженной возбудимости (тетанус – минимален).
При изучении скелетных и сердечных мышц в поляризованном свете, видны чередующиеся зоны с различной оптической плотностью (рис. 3). Это позволило разделить сократительные элементы поперечнополосатых мышечных волокон (миофибрилл) на функциональные единицы — саркомеры (участки между соседними Z-мембранами).
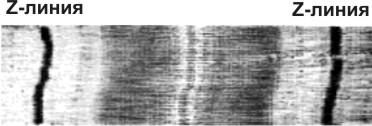
Рис. 3. Микрофотография участка поперечнополосатой мышцы.
Характерная оптическая плотность участков саркомера обусловлена особенностями расположения в них (рис. 4) сократительных белков (актина и миозина).

Рис. 4. Схема расположения сократительных белков в саркомере (в Н-зоне миозиновые фибриллы не имеют мостиков).
Около Z-мембран содержится актин (светлая область – изотропный или I-диск). Ближе к середине саркомера видны темные (анизотропные) А-диски с миозином и актином. В центре саркомера проходит М-линия. По обе стороны от нее расположен миозин (просветление или Н-зона). Актин с одной стороны прикреплен к Z-мембране, а с другой свободен и заканчивается между миозиновыми волокнами в области А-диска (на границе с Н-зоной). Оба конца миозиновой нити свободны.
При сокращении уменьшается ширина только изотропных дисков. Миозиновые нити при этом могут достигать своими концами Z-пластинок, а длина саркомеров — укорачиваться на 30-50%.
Механизм мышечного сокращения. Практически на всей миозиновой нити имеются боковые мостики (отсутствуют только около М-линии). После сцепления с актином они изменяют угол наклона (используя энергию АТФ), что продвигает сократительные белки относительно друг друга (саркомер укорачивается). Затем актин соединяется с другим мостиком миозина и совершается дальнейшее продвижение.
В покое соединению актина с миозином мешают белки тропонин и тропомиозин. При возбуждении они «нейтрализуются» поступающим в саркоплазму (цитоплазму мышечной клетки) кальцием и начинается взаимодействие сократительных белков. Прекращение возбуждения приводит к удалению кальция из саркоплазмы, тропонин с тропомиозином разрушают комплекс актина и миозина – мышца расслабляется.
В скелетных мышцах используется кальций из внутриклеточных цистерн саркоплазматической сети. Гладкие мышцы получают кальций только из межклеточных пространств, а сердечная мускулатура использует оба источника данного иона. Использование только внутриклеточных источников кальция позволяет скелетным мышцам сокращаться и расслабляться с наибольшей скоростью, а гладкая мускулатура — медленно изменяет свой тонус.
Синапс — функциональное соединение между нейроном и другими клетками. Существуют электрические и химические синапсы. Электрическим синапсам свойственно низкое электрическое сопротивление в области контакта клеток и ПД быстро передается на соседнюю мембрану. Химические синапсы состоят из пресинаптической мембраны, постсинаптической мембраны и синаптической щели (рис. 5.). Под влиянием нервных импульсов, в химических синапсах происходит высвобождение химического вещества — медиатора (например, ацетилхолина, норадреналина) из пресинаптических утолщений аксона в синаптическую щель и его взаимодействие с рецепторами на постсинаптической мембране. В возбуждающих синапсах это приводит к возникновению возбуждающего (ВПСП), а в тормозных — тормозного (ТПСП) постсинаптических потенциалов. После того как медиатор выполнил свою функцию, он разрушается или поглощается клетками.
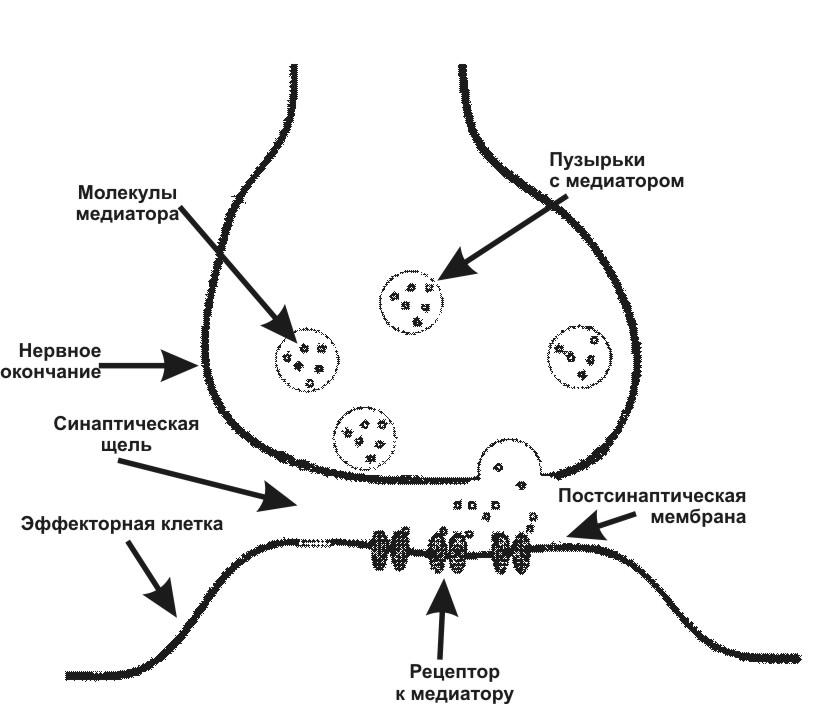
Рис. 5. Схема строения химического синапса.
За пределами ЦНС наиболее распространен медиатор ацетилхолин. Он облегчает проведение возбуждения через вегетативные ганглии, повышает секрецию адреналина надпочечниками и соляной кислоты желудочными железами, угнетает работу сердца, вызывает сокращение гладких мышц некоторых внутренних органов и экзокринных желез. В гладких мышцах бронхов, кишечника, мочевого пузыря, матки, круговой и цилиарной мышцах глаза ацетилхолин приводит, соответственно, к бронхоспазму, усилению перистальтики кишечника и желудка (при расслаблении сфинктеров), сокращению мочевого пузыря и сужению зрачка.
По расположению синапсы делят на периферические (нервно-мышечные, рецепторно-нейрональные и т.д.); центральные (аксосоматические, аксодендритные и др.); по знаку действия (возбуждающие и тормозящие) и по выделяемому медиатору (холинергические, адренергические и др.).
Химические синапсы проводят возбуждение в одном направлении; передают возбуждение медленнее, чем по нервному волокну (синаптическая задержка); имеют низкую лабильность, а также высокую утомляемость и чувствительность к химическим веществам.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом