
В каких тканях наиболее интенсивны реакции пентозофосфатного пути
Открытие пути прямого окисления углеводов, или, как его называют, пентозного пути, связано главным образом с работами Варбурга, Липмана, Диккенса и В. А. Энгельгардта. Расхождение путей окисления углеводов — классического, при участии цикла трикарбоновых кислот (цикла Кребса), и пентозного — начинается со стадии образования гексозомонофосфата. Если глюкозо-6-фосфат (точнее, фруктозо-6-фосфат) подвергается еще раз фосфорилированию и превращается во фруктозо-1,6-дифосфат, то в этом случае дальнейший распад углеводов происходит по обычному гликолитическому пути с образованием пировиноградной кислоты, которая, окиcляясь до ацетил-КоА, затем сгорает в цикле Кребса.
Если же присоединения второй частицы фосфата к гексозо-6-монофосфорному эфиру не происходит, то фосфорилированная глюкоза может подвергаться прямому окислению до фосфопентоз. В норме доля пентозного цикла в количественном превращении глюкозы обычно невелика, варьирует у разных организмов и зависит от типа ткани и ее функционального состояния.
У млекопитающих активность пентозного цикла относительно высока в печени, надпочечниках, в эмбриональной ткани и в молочной железе в период лактации. Значение этого пути в обмене веществ велико. Он поставляет восстановленный НАДФ (НАДФН2), необходимый для биосинтеза жирных кислот, холестерина и т. д. За счет пентозного цикла примерно на 50% покрывается потребность организма в НАДФН2.

Вторая функция пентозного цикла заключается в том, что он поставляет пентозофосфаты для синтеза нуклеиновых кислот и многих коферментов. При ряде патологических состояний удельный вес пентозного пути окисления глюкозы возрастает. Механизм реакций пентозного цикла достаточно расшифрован. Пентозный цикл начинается с окислительного декарбоксилирования (от гексозофосфата отщепляется первый атом углерода). Это так называемая окислительная стадия пентозного цикла. Вторая стадия включает неокислительные превращения пентозофосфатов с образованием исходного глюкозо-6-фосфата (рис. 93).
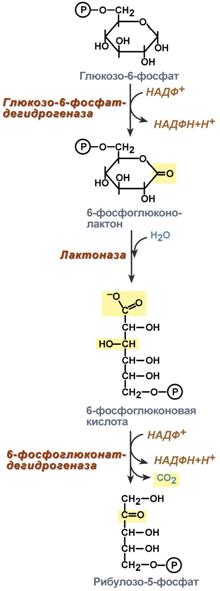
Реакции пентозного цикла протекают в цитоплазме клетки. Первая реакция — дегидрирование глюкозо-6-фосфата при участии фермента глюкозо-6-фосфат-дегидрогеназы и кофермента НАДФ:

Образовавшийся в ходе данной реакции 6-фосфоглюконой-σ-лактон — соединение нестабильное и с большой скоростью гидролизуется либо спонтанно, либо с помощью фермента глюконо-лактоназы:

Равновесие первой суммарной реакции сильно смещено в сторону образования НАДФН2.
В следующей окислительной реакции, катализируемой фосфоглюконатдегидрогеназой (декарбоксилирующей), 6-фосфоглюконовая кислота дегидрируется и декарбоксилируется. В результате образуется фосфорилированная кетопентоза-D-рибулозо-5-фосфат и еще одна молекула НАДФН2. В качестве промежуточного продукта в реакции, вероятно, образуется 3-кето-6-фосфоглюконовая кислота:

Под действием соответствующей эпимеразы из рибулозо-5-фосфата может образоваться другая фосфопентоза — ксилулозо-5-фосфат. Кроме того, рибулозо-5-фосфат под влиянием особой изомеразы легко превращается в рибозо-5-фосфат. Между этими формами пентозофосфатов устанавливается состояние подвижного равновесия:

При определенных условиях пентозный путь на этом этапе может быть завершен. Суммарный итог окислительной стадии (этапа) можно выразить в виде следующего уравнения:
Глюкозо-6-фосфат + 2НАДФ —> Пентозо-5-фосфат + СO2 + 2НАДФН2
Однако при других условиях наступает так называемый неокислительный этап (стадия) пентозного цикла. Реакции этого этапа не связаны с использованием кислорода и протекают в анаэробных условиях. При этом частично образуются вещества, характерные для первой стадии гликолиза (фруктозо-6-фосфат, фруктозо-1,6-дифосфат, фосфотриозы), а частично — специфические для пентозного пути (седогептулозо-7-фосфат, пентозо-5-фосфат, эритрозо-4-фосфат).
Основными реакциями неокислительной стадии пентозного цикла являются транскетолазная и трансальдолазная. Эти реакции катализируют превращение изомерных пентозо-5-фосфатов:

Коферментом в транскетолазной реакции служит тиамин-дифосфат (ТДФ), играющий роль промежуточного переносчика гликольальдегидной группы, которая затем присоединяется к рибозо-5-фосфату. В результате образуется семиуглеродный сахар — седогептулозо-7-фосфат и глицеральдегид-3-фосфат.
Транскетолазная реакция в пентозном цикле встречается дважды, второй раз — при образовании фруктозо-6-фосфата и триозофосфата в результате взаимодействия второй молекулы ксилулозо-5-фосфата с эритрозо-4-фосфатом:

Фермент трансальдолаза катализирует перенос остатка дигидроксиацетона (но не свободного дигидроксиацетона) от седогептулозо-7-фосфата на глицеральдегид-3-фосфат:

Как видно из рис. 93, 6 молекул глюкозо-6-фосфата, вступая в пентозный цикл, дают 6 молекул рибулозо-5-фосфата и 6 молекул СО2, после чего из 6 молекул рибулозо-5-фосфата снова регенерируют 5 молекул глюкозо-6-фосфата. При этом молекула глюкозо-6-фосфата, вступающая в цикл, полностью не окисляется. Шесть молекул СО2 образуются из C1-групп 6 молекул глюкозо-6-фосфата. Валовое уравнение окислительной и неокислительной стадий пентозного цикла можно представить в следующем виде:
Читайте также: Представляют собой мельчайшие микроорганизмы поражающие ткани человека
6 Глюкозо-6-фосфат + 7 Н2O + 12 НАДФ —> 5 Глюкозо-6-фосфат+ + 6 СО2 + Фн + 12 НАДФН,
или
Глюкозо-6-фосфат + 7 Н2О + 12 НАДФ —> 6СO2 + Н3РO4 + 12 НАДФН2

Образовавшийся НАДФН2 используется на восстановительные синтезы в цитоплазме и, как правило, не участвует в окислительном фосфорилировании, протекающем в митохондриях.
В последние годы появились работы, которые дают основание предполагать, что схема пентозного превращения углеводов сложнее, чем это представлено на рис. 93.
Согласно новой схеме пентозофосфатного пути, предложенной Вильямсом и Кларком, первые этапы превращения совпадают с прежней схемой. Однако после первой транскетолазной реакции сразу наступает отклонение: рибозо-5-фосфат (Р-5-Ф) под влиянием пентозофосфатэпимеразы (ПФЭ) превращается в арабинозо-5-фосфат (Араб-5-Ф); а глицеральдегид-3-фосфат (Га-3-Ф) изомеризуется в дигидроксиацетонфосфат (ДАФ). Между этими веществами происходит реакция при участии альдолазы и образуется октулозо-1,8-дифосфат (0-1,8-диФ). Что же касается второго продукта транскетолазной реакции — седогептулозо-7-фосфата (С-7-Ф), то он переходит под влиянием фосфотрансферазы в седогептулозо-1,7-дифосфат (С-1,7-диФ) с одновременным образованием октулозо-8-фосфата (0-8-Ф). Затем наступает вторая альдолазная реакция, которая приводит к превращению седогептулозо-1,7-дифосфата в эритрозо-4-фосфат (Э-4-Ф) и дигидрооксиацетонфосфат. Далее эритозо-4-фосфат и октулозо-8-фосфат в результате второй реакции образуют фруктозо-6-фосфат (Ф-6-Ф) и глюкозо-6-фосфат (Гл-6-Ф). Все приведенные на рис. 94 реакции обратимы.
Многие исследователи считают, что пентозный путь (его анаэробная часть) и гликолиз, протекающие в цитолизе, способны переключаться с одного пути на другой в зависимости от соотношения концентраций промежуточных продуктов, образовавшихся в клетке. Итак, следует признать возможным обобщение в один суммарный процесс анаэробной фазы пентозного пути превращения углеводов и гликолиза. При этом роль важнейшего регулятора данного процесса играет эритрозо-4-фосфат. В зависимости от того, происходит ли интенсивное использование фосфопентоз или фосфопентозы образуются в оптимальном избытке, эритрозо-4-фосфат участвует либо в альдолазной реакции с образованием седогептулозо-1,7-дифосфата, либо в транскетолазной реакции с образованием фруктозо-6-фосфата и глюкозо-6-фосфата.
Для синтеза рибозы и НАДФН существует специальный путь
Так как в клетках непрерывно просходят реакции синтеза белков, то для этого процесса требуются рибонуклеиновые кислоты. В свою очередь для синтеза самих нуклеиновых кислот, а точнее пуриновых и пиримидиновых нуклеотидов, требуется рибозо-5-фосфат . Если клетка готовится к делению, то для синтеза ДНК ей нужны дезоксирибонуклеотиды, которые образуются при участии НАДФН .
Молекулы НАДФН также используются:
- для синтезажирных кислот (печень, жировая ткань),
- для синтезахолестерола и других стероидов (например, желчных кислот),
- в реакциия восстановительного аминирования для образованияглутаминовой кислоты из α-кетоглутаровой кислоты,
- для восстановления окисленного тиоредоксина и окисленного глутатиона в системе антиоксидантной защиты клетки от свободно-радикального окисления,
- для восстановления железа (III) в метгемоглобине в железо (II) нормального гемоглобина (эритроциты),
- для реактивации фолиевой кислоты и витамина К.
В клетке существует процесс, обеспечивающий одновременное образование рибозы и НАДФН – это пентозофосфатный путь .
Пентозофосфатный путь
Наиболее активно реакции пентозофосфатного пути идут в цитозоле клеток печени, жировой ткани, эритроцитах, коре надпочечников, молочной железе при лактации, в гораздо меньшей степени в скелетных мышцах. Этот путь окисления глюкозы не связан с образованием энергии, а обеспечивает анаболизм клеток. В связи с этим у новорожденных и детей первых лет жизни его активность довольно высока.
Пентозофосфатный путь включает два этапа – окислительный и этап структурных перестроек (неокислительный).
Первый этап
На этом этапе происходит регуляция процесса: инсулин повышает активность глюкозо-6-фосфат-дегидрогеназы и фосфоглюконат-дегидрогеназы.
Реакции первого этапа
Второй этап
Второй этап – этап структурных перестроек , благодаря которым пентозы способны возвращаться в фонд гексоз.
В этих реакциях молекулы рибулозо-5-фосфата изомеризуются до рибозо-5-фосфата и ксилулозо-5-фосфата. Далее под влиянием ферментов транскетолазы и трансальдолазы происходят структурные перестройки с образованием других моносахаридов.
При реализации всех реакций второго этапа пентозы превращаются во фруктозо-6-фосфат и глицеральдегид-3-фосфат, т.е. возвращаются в реакции гликолиза. Поэтому иногда пентозофосфатный путь называют шунтом, т.е. рассматривают как обходной путь окисления глюкозы.
Глицеральдегид-3-фосфат в зависимости от условий и вида клеток может
Биохимия метаболизма. Учебное пособие

Учебник описывает последовательность и механизмы реакций основных метаболитических путей энергетического и пластического обмена. Описываются ингибиторы реакций и механизмы их действия, регуляция скорости метаболитических путей. В разработке использовались как классические данные о метаболитических путях, так и новая информация об механизмах реакций и их регуляции. Учебник предназначен для студентов любых ВУЗов для изучения биохимии, в качестве как основной, так и дополнительной литературы.
Оглавление
Приведённый ознакомительный фрагмент книги Биохимия метаболизма. Учебное пособие предоставлен нашим книжным партнёром — компанией ЛитРес.
Пентозофосфатный путь является альтернативным путем окисления глюкозы. Он включает несколько этапов, в результате функционирования которых из трех молекул глюкоза-6-фосфата образуются три молекулы СО2 и три молекулы пентоз. Последние используются для регенерации двух молекул глюкозо-6-фосфата и одной молекулы глицеральдегид-3-фосфата. Поскольку из двух молекул глицеральдегид-3-фосфата можно регенерировать молекулу глюкоза-б-фосфата, глюкоза может быть полностью окислена при превращении по пентозофосфатному пути.
У растений часть реакций пентозофосфатного пути участвует также в образовании гексоз из СО2 при фотосинтезе. Пентозофосфатный путь называют иногда пентозным шунтом, гексозомонофосфатным путем или фосфоглюконатным окислительным путем. Открытие Отто Варбургом (Otto Warburg) в 1931 г. глюкозо-6-фосфат-дегидрогеназы, первого фермента этого пути, сделало возможной его полную расшифровку, которую осуществили Фриц Липман, Фрэнк Дикенс, Бернард Хорекер и Эфроим Рэкер.
Пентозофосфатный цикл не приводит к синтезу АТР, он выполняет две главные функции: 1) образование NADPH для восстановительных синтезов, таких, как синтез жирных кислот и стероидов; 2) обеспечение рибозой синтеза нуклеотидов и нуклеиновых кислот. Ферменты пентозофосфатного пути локализованы во внемитохондриальном пространстве клеткив цитозоле. Как и в процессе гликолиза, окисление осуществляется путем дегидрогенирования, однако акцептором водорода в этом случае служит не NAD, а NADP.

Рисунок 4: Схема реакций пентозофосфатного шунта (черный круг с буквой Р в центре обозначает фосфатную группу).
Последовательность реакций пути можно разделить на две фазы: окислительную и неокислительную (схема реакций представлена на рисунке 4). В реакциях первой фазы глюкоза-6-фосфат дегидрогенируется и декарбоксилируется с образованием рибулозо-5-фосфата. В ходе второй фазы рибулозо-5-фосфат превращается снова в глюкозо-6-фосфат в результате серии реакций, в которых главную роль играют два фермента: транскетолаза и трансальдолаза
Окислительная фаза пентозофосфатного пути начинается с дегидрирования глюкозо-6-фосфата при С-1, реакции, катализируемой глюкозо-6-фосфат-дегидрогеназой.
Фермент высокоспецифичен в отношении NADP + ; Км для NAD + примерно в тысячу раз выше, чем для NADP + . Продуктом реакции является 6-фосфоглюконо-δ-лактон, внутримолекулярный эфир, с эфирной связью между С-1-карбоксильной группой и гидроксилом при С-5. Следующий этап — гидролиз 6-фосфоглюконо — δ — лактона специфической лактоназой, дающий 6-фосфо-глюконат. Этот шестиуглеродный сахар подвергается затем окислительному декарбоксилированию 6-фосфоглюконат — дегидрогеназой с образованием рибулозо-5-фосфата. Акцептором электронов вновь служит NADP + . (смотри рисунок). Конечным этапом синтеза рибозо-5-фосфата является изомеризация рибулозо-5-фосфата фосфопентозо-изомеразой. Эта реакция подобна гликолитическим реакциям:
Все три кетозо-альдозные изомеризации идут через образование ендиольного промежуточного продукта.
Окислительная фаза пентозофосфатного пути иногда считается основной и неокислительная рассматривается как связка с гликолизом. Это связано с тем фактом, что эти фазы могут идти как независимо друг от друга, так вместе. В ходе неокислительной фазы пентозофосфатного пути происходит регенерация рибозо-5-фосфата в глюкозо-6-фосфат. Основную роль в этом процессе играют два фермента: транскетолаза и трансалъдолаза. Эти же ферменты создают обратимую связь между пентозофосфатным путем и гликолизом, катализируя следующие три реакции:
Транскетолаза переносит двухуглеродную группу, включающую 1-й и 2-й атомы углерода кетозы, на альдегидный углерод альдозного сахара. Происходит, следовательно, превращение кетосахара в альдозу, содержащую на два атома углерода меньше, и одновременное превращение альдосахара в кетозу, содержащую на два атома углерода больше. Коферментом реакции является тиаминидифосфат (в его состав входит тиамин — витамин группы В), для ее протекания необходимы ионы Mg 2+ . Переносимая двухуглеродная группа является, вероятно, гликоальдегидом, связанным с тиаминдифосфатом, т. е. «активным гликольальдегидом». Транскетолаза катализирует перенос двухуглеродной группы с ксилулозо-5-фосфата на рибозо-5-фосфат с образованием семиуглеродной кетозы седогептулозо-7-фосфата и альдозы глицеральдегид-3-фосфата. Эти два продукта далее вступают в следующую реакцию, называемую трансальдолазной. Трансальдолаза осуществляет перенос трехуглеродного фрагмента, «активного дигидроксиацетона» (атомы углерода 1 — 3), кетозы седогептулозо-7-фосфата на альдозу глицеральдегид-3-фосфат; в результате образуются кетоза фруктоза-6-фосфат и четырехуглеродная альдоза эритрозо-4-фосфат. Следующая реакция снова катализируется транскетолазой. В этой реакции ксилулозо-5-фосфат служит донором «активного гликоальдегида». Роль акцептора выполняет образовавшийся ранее эритрозо-4-фосфат. Продуктами этой реакции являются фруктоза-6-фосфат и глицеральдегид-3-фосфат.
Итак, избыток рибозо-5-фосфата, образованный в пентозофосфатном пути, может количественно превращаться в промежуточные продукты гликолиза.
Значение метаболического пути для различных тканей можно оценить по его активности. Пентозофосфатный путь активно протекает в печени, жировой ткани, коре надпочечников, щитовидной железе, эритроцитах, семенниках и в молочных железах в период лактации; он неактивен в нелактирующей молочной железе и малоактивен в скелетных мышцах. Все ткани, в которых активность данного пути высока, используют в реакциях восстановительного синтеза NADPH, например в реакциях синтеза жирных кислот, стероидов, аминокислот (с участием глутаматдегидрогеназы) или восстановленного глутатиона в эритроцитах. Вероятно, в условиях активного липогенеза или при наличии любой системы, утилизирующей NADPH, возрастает активная деградация глюкозы по пентозофосфатному пути в связи с увеличением отношения NADP + /NADPH. В условиях, которые возникают после приема пищи, может индуцироваться синтез глюкоза-6-фосфатдегидрогеназы и 6-фосфоглюконатдегидрогеназы.
Регуляция скорости функционирования пентозофосфатного пути
Первая реакция окислительной ветви пентозофосфатного пути, дегидрирование глюкозо-6-фосфата, по существу необратима. Действительно, при физиологических условиях эта реакция лимитирует скорость процесса и выполняет функцию «контрольного пункта». Наиболее важным регуляторным фактором является концентрация NADP + , акцептора электронов при окислении глюкозо-6-фосфата в 6-фосфоглюконо-лактон. Кроме того, NADPH конкурирует с NADP + за связывание с ферментом, и АТР конкурирует с глюкозо-6-фосфатом. Отношение концентрации NADP + к концентрации NADPH в цитозоле печени крыс, содержащихся на полноценном рационе, составляет примерно 0,014, что на несколько порядков ниже отношения [NAD + ] / [NADH], которое при этих же условиях равно 700. Выраженное действие концентрации NADP + на скорость превращений по окислительной ветви пентозофосфатного пути подтверждает, что генерирование NADPH тесно сопряжено с его использованием в восстановительных биосинтезах. Вопрос о регуляции неокислительной ветви пентозофосфатного пути до сих пор остается открытым.
Регуляция направления пентозофосфатного шунта
Судьба глюкозо-6-фосфата зависит от потребности в NADPH, рибозо-5-фосфате и АТФ.
В данном случае возможно четыре различные ситуации (схема регуляторных механизмов представлена на рисунке 5).

Рисунок 5: Схема регуляции направлений пентозофосфатного шунта
1. Потребность в рибозо-5-фосфате значительно превышает потребность в NADPH. Большая часть глюкозо-6-фос-фата превращается во фруктозо-6-фостфат и глицеральдегид-3-фосфат по гликолитическому пути. Затем две молекулы фрукто-зо-6-фосфата и одна молекула глицераль-дегид-3-фосфата превращаются под действием трансальдолазы и транскетолазы в три молекулы рибозо-5-фосфата путем обращения реакции, описанной ранее. Стехиометрия этого превращения следующая:
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом