
Пентозофосфатный шунт (путь, цикл) является альтернативным путем окисления глюкозы. Наиболее активен этот процесс в жировой ткани, печени, коре надпочечников, эритроцитах, фагоцитирующих лейкоцитах, лактирующей молочной железе, семенниках. Протекает он в цитозоле без участия кислорода и состоит из 2 стадий окислительной и неокислительной. В окислительной стадии происходит восстановление НАДФН2, который используется: 1) для регенерации глутатиона в антиоксидантной системе; 2) для синтеза жирных кислот; 3) в оксигеназных реакциях с участием цитохрома Р450 при обезвреживании ксенобиотиков, метаболитов, синтезе холестерина, стероидных гормонов и т.д. В неокислительной стадии образуются различные пентозы. Рибозо-5ф может использоваться для синтеза пуриновых и пиримидиновых нуклеотидов.
Тканевые особенности функционирования ПФШ (пути, цикла).
В зависимости от потребности ткани, пентозофосфатный процесс может протекать в виде метаболического цикла, пути или шунта начальных реакций гликолиза:
1. При ПФЦ или ПФШ в качестве продукта образуется только НАДФН2. Пентозы в этом случае не являются конечным продуктом, они превращаются в фосфогексозы, которые замыкают цикл, или уходят в гликолиз, завершая шунт. В жировой ткани, эритроцитах.
2. Продуктом ПФП являются НАДФН2 и пентозы. В печени, костном мозге.
3. В тканях, которые не испытывают потребность в НАДФН2, функционирует только неокислительная стадия ПФП, причем ее реакции идут в обратную сторону начиная с фруктозы-6ф до фосфопентоз. В мышцах.
Реакции окислительной стадии
Окислительная стадия ПФШ (пути, цикла) состоит из 3 необратимых реакций:
1). Глюкозо-6ф дегидрогеназа (глюкозо-6ф: НАДФ + оксидоредуктаза). Ингибитор НАДФН2. Индуктор инсулин.
2). Глюконолактонгидратаза (6-фосфоглюконат: гидро-лиаза).
3). 6-фосфоглюконат дегидрогеназа (6-фосфоглюконат: НАДФ + оксидоредуктаза (декарбоксилирующая)). Индуктор инсулин.
Схема ПФШ (пути, цикла)
На схеме неокислительная стадия начинается с эпимераз и изомераз, которые изомеризуют рибулозо-5ф. Все реакции неокислительной стадии обратимы.
Общее уравнение ПФЦ:
6 глюкозо-6ф + 12 НАДФ + → 6 СО2 + 12 НАДФН2 + 5 глюкозо-6ф
Общее уравнение ПФШ:
3 глюкозо-6ф + 6 НАДФ + → 3 СО2 + 6 НАДФН2 + 2 фруктозо-6ф + ФГА
Общие уравнения ПФП:
1) глюкозо-6ф + 2 НАДФ + → СО2 + 2 НАДФН2 + рибозо-5ф
2) 2 фруктозо-6ф + ФГА → 3 рибозо-5ф
Патология ПФШ
НАДФН2 является важным компонентом антиоксидантной защиты, он необходим для регенерации глутатиона, который с участием глутатионпероксидазы разрушает активные формы кислорода. Так как в эритроцитах НАДФН2 образуется только в реакциях ПФШ, дефект глюкозо-6ф ДГ вызывает дефицит НАДФН2 и снижение антиоксидантной защиты. В этом случае под действием прооксидантов, например, антималярийных препаратов происходит существенное повышение СРО. Активация СРО вызывает окисление цистеина в белковой части гемоглобина, в результате чего протомеры гемоглобина, соединяясь дисульфидными мостиками, образуют тельца Хайнца. Т.к. тельца Хайнца снижают пластичность клеточной мембраны эритроцитов, она при деформации в капиллярах разрушается. Массированный гемолиз эритроцитов ведет к развитию гемолитической анемии.
Витамин B1 (тиамин).
Структура витамина включает пиримидиновое и тиазоловое кольца, соединённые метановым мостиком.
Источники. Витамин В1 — первый витамин, выделенный в кристаллическом виде К. Фун-ком в 1912 г. Он широко распространён в продуктах растительного происхождения (оболочка семян хлебных злаков и риса, горох, фасоль, соя и др.). В организмах животных витамин В1, содержится преимущественно в виде дифосфорного эфира тиамина (ТДФ); он образуется в печени, почках, мозге, сердечной мышце путём фосфорилирования тиамина при участии тиаминкиназы и АТФ.
Суточная потребность взрослого человека в среднем составляет 2-3 мг витамина В1. Но потребность в нём в очень большой степени зависит от состава и общей калорийности пищи, интенсивности обмена веществ и интенсивности работы. Преобладание углеводов в пище повышает потребность организма в витамине; жиры, наоборот, резко уменьшают эту потребность.
Читайте также: Ткани для кимоно какие
Биологическая роль витамина В1, определяется тем, что в виде ТДФ он входит в состав как минимум трёх ферментов и ферментных комплексов: в составе пируват- и α-кетоглутаратдегидрогеназных комплексов он участвует в окислительном декарбокси-лировании пирувата и α-кетоглутарата; в составе транскетолазы ТДФ участвует в пентозофосфатном пути превращения углеводов.
Основной, наиболее характерный и специфический признак недостаточности витамина В1 — полиневрит, в основе которого лежат дегенеративные изменения нервов. Вначале развивается болезненность вдоль нервных стволов, затем — потеря кожной чувствительности и наступает паралич (бери-бери). Второй важнейший признак заболевания — нарушение сердечной деятельности, что выражается в нарушении сердечного ритма, увеличении размеров сердца и в появлении болей в области сердца. К характерным признакам заболевания, связанного с недостаточностью витамина В1, относят также нарушения секреторной и моторной функций ЖКТ; наблюдают снижение кислотности желудочного сока, потерю аппетита, атонию кишечника.
ПЕНТОЗОФОСФАТНЫЙ ПУТЬ ОКИСЛЕНИЯ УГЛЕВОДОВ
Открытие пути прямого окисления углеводов, или, как его называют, пентозофосфатного цикла, принадлежит О. Варбургу, Ф. Липману, Ф. Дикенсу и В.А. Энгельгарду. Расхождение путей окисления углеводов – классического (цикл трикарбоновых кислот, или цикл Кребса) и пентозофосфатного – начинается со стадии образования гексозомонофосфата. Если глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат, который фосфорилируется второй раз и превращается во фруктозо-1,6-бисфосфат, то в этом случае дальнейший распад углеводов происходит по обычному гликолитическому пути с образованием пировиноградной кислоты, которая, окисляясь до ацетил-КоА, затем «сгорает» в цикле Кребса.
Если второго фосфорилирования гексозо-6-монофосфата не происходит, то фосфорилированная глюкоза может подвергаться прямому окислению до фосфопентоз. В норме доля пентозофосфатного пути в количественном превращении глюкозы обычно невелика, варьирует у разных организмов и зависит от типа ткани и ее функционального состояния.
У млекопитающих активность пентозофосфатного цикла относительно высока в печени, надпочечниках, эмбриональной ткани и молочной железе в период лактации. Значение этого пути в обмене веществ велико. Он поставляет восстановленный НАДФН, необходимый для биосинтеза жирных кислот, холестерина и т.д. За счет пентозофосфатного цикла примерно на 50% покрывается потребность организма в НАДФН.
Другая функция пентозофосфатного цикла заключается в том, что он поставляет пентозофосфаты для синтеза нуклеиновых кислот и многих коферментов. При ряде патологических состояний удельный вес пенто-зофосфатного пути окисления глюкозы возрастает. Механизм реакций пентозофосфатного цикла достаточно расшифрован.
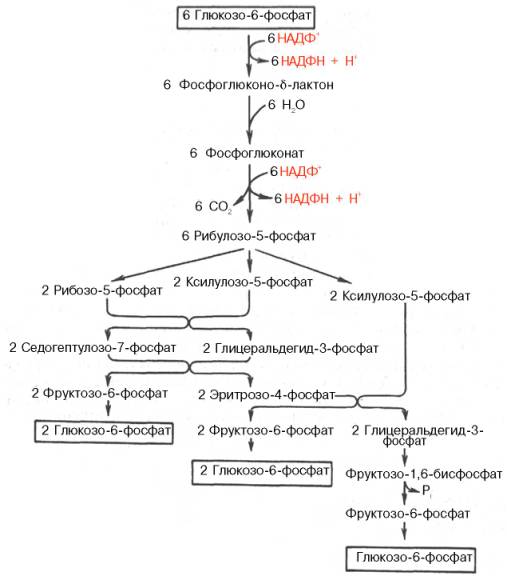
Рис. 10.12. Пентозофосфатный путь окисления углеводов.
Пентозофосфатный цикл начинается с окисления глюкозо-6-фосфата и последующего окислительного декарбоксилирования продукта (в результате от гексозофосфата отщепляется первый атом углерода). Это первая, так называемая окислительная, стадия пентозофосфатного цикла. Вторая стадия включает неокислительные превращения пентозофосфатов с образованием исходного глюкозо-6-фосфата (рис. 10.12). Реакции пен-тозофосфатного цикла протекают в цитозоле клетки.
Первая реакция – дегидрирование глюкозо-6-фосфата при участии фермента глюкозо-6-фосфатдегидрогеназы и кофермента НАДФ + . Образовавшийся в ходе реакции 6-фосфоглюконо-δ-лактон – соединение нестабильное и с большой скоростью гидролизуется либо спонтанно, либо с помощью фермента 6-фосфоглюконолактоназы с образованием 6-фос-фоглюконовой кислоты (6-фосфоглюконат):
Читайте также: Оао витебский комбинат шелковых тканей
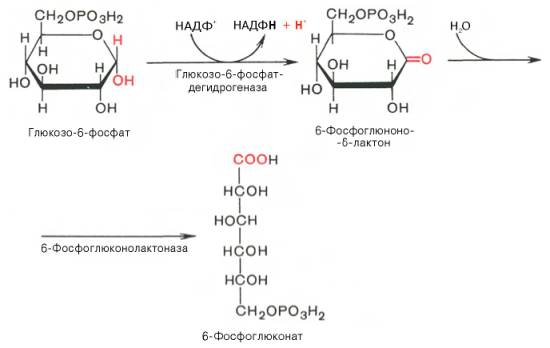
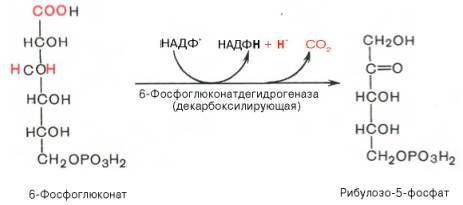
Во второй – окислительной – реакции, катализируемой 6-фосфоглюко-натдегидрогеназой (декарбоксилирующей), 6-фосфоглюконат дегидрируется и декарбоксилируется. В результате образуется фосфорилированная кетопентоза – D-рибулозо-5-фосфат и еще 1 молекула НАДФН:
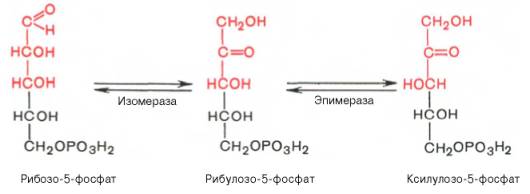
Под действием соответствующей эпимеразы из рибулозо-5-фосфата может образоваться другая фосфопентоза – ксилулозо-5-фосфат. Кроме того, рибулозо-5-фосфат под влиянием особой изомеразы легко превращается в рибозо-5-фосфат. Между этими формами пентозофосфатов устанавливается состояние подвижного равновесия:
При определенных условиях пентозофосфатный путь на этом этапе может быть завершен. Однако при других условиях наступает так называемый неокислительный этап (стадия) пентозофосфатного цикла. Реакции этого этапа не связаны с использованием кислорода и протекают в анаэробных условиях. При этом образуются вещества, характерные для первой стадии гликолиза (фруктозо-6-фосфат, фруктозо-1,6-бисфосфат, фосфотрио-зы), а другие – специфические для пентозофосфатного пути (седогептулозо-7-фосфат, пентозо-5-фосфаты, эритрозо-4-фосфат).
Основными реакциями неокислительной стадии пентозофосфатного цикла являются транскетолазная и трансальдолазная. Эти реакции катализируют превращение изомерных пентозо-5-фосфатов:
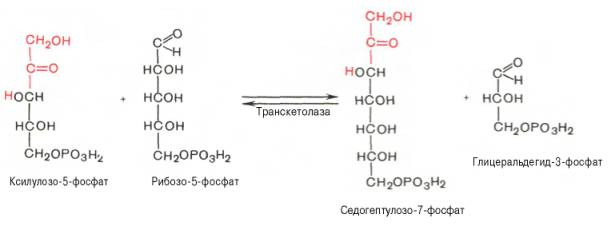
Коферментом в транскетолазной реакции служит ТПФ, играющий роль промежуточного переносчика гликольальдегидной группы от ксилулозо-5-фосфата к рибозо-5-фосфату. В результате образуется семиуглеродный моносахарид седогептулозо-7-фосфат и глицеральдегид-3-фосфат.
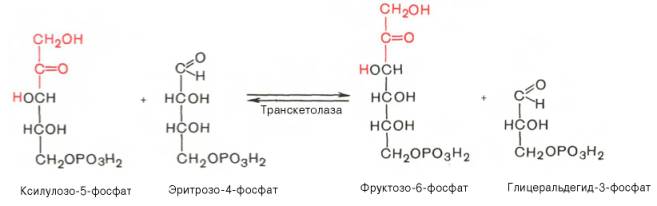
Транскетолазная реакция в пентозном цикле встречается дважды, второй раз – при образовании фруктозо-6-фосфата и триозофосфата в результате взаимодействия второй молекулы ксилулозо-5-фосфата с эритрозо-4-фосфатом:
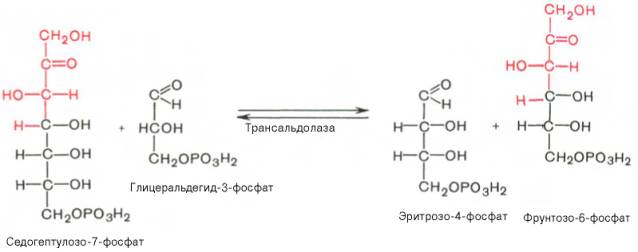
Фермент трансальдолаза катализирует перенос остатка диоксиацетона (но не свободного диоксиацетона) от седогептулозо-7-фосфата на гли-церальдегид-3-фосфат:
Шесть молекул глюкозо-6-фосфата, вступая в пентозофосфатный цикл, образуют 6 молекул рибулозо-5-фосфата и 6 молекул СО2, после чего из 6 молекул рибулозо-5-фосфата снова регенерируется 5 молекул глюко-зо-6-фосфата (см. рис. 10.12). Однако это не означает, что молекула глюкозо-6-фосфата, вступающая в цикл, полностью окисляется. Все 6 молекул СО2 образуются из С-1-атомов 6 молекул глюкозо-6-фосфата.
Валовое уравнение окислительной и неокислительной стадий пенто-зофосфатного цикла можно представить в следующем виде:

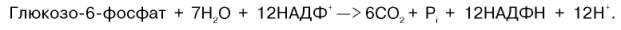
Образовавшийся НАДФН используется в цитозоле на восстановительные синтезы и, как правило, не участвует в окислительном фосфори-лировании, протекающем в митохондриях.
В последние годы появились работы, которые дают основание предполагать, что в некоторых тканях схема пентозофосфатного превращения углеводов сложнее, чем это представлено на рис. 10.12. Согласно этой более полной схеме пентозофосфатного пути, первые этапы превращения совпадают с прежней схемой, однако после первой транскетолазной реакции начинаются некоторые отклонения (рис. 10.13).
Считают, что пентозофосфатный путь и гликолиз, протекающие в ци-тозоле, взаимосвязаны и способны переключаться друг на друга в зависимости от соотношения концентраций промежуточных продуктов, образовавшихся в клетке (см. рис. 10.13).
ПЕНТОЗОФОСФАТНЫЙ ПУТЬ
Пентозофосфатный путь (ПФП) обмена углеводов нередко называют апотомическим путём, так как обмен глюкозы идёт по первому (С1) атому углерода.
Доля ПФП в количественном превращении глюкозы в клетках обычно невелика (в большинстве клеток не более 10 %) и варьирует в зависимости от типа ткани и её функ-ционального состояния. Так, в клетках печени по этому пути превращается до 20 % глюкозы, в эритроцитах — 7 %, в клетках мозга — около 2 %. Этот процесс идет в клетках многих органов и тканей.
Ферменты ПФП локализованы в цитоплазме клеток.
Превращение глюкозы по ПФП не требует присутствия кислорода. Если по ПФП превращается шесть молекул Гл-6-Ф, то за один цикл молекула Гл-6-Ф катаболизирует до 6 СО2.
6 Гл-6-Ф + 7H2O + 12 НАДФ + ® 5 Гл-6-Ф + 6СО2 + 12 НАДФН . Н + + ФН
Читайте также: Ткань водопоглощающая aion plas senu в тубе 69х43 см голубая
Последовательность реакций ПФП разделяют на два этапа:
I. Окислительный этап. На этом этапе осуществляются две дегидрогеназные реакции и одна реакция декарбоксилирования с образованием рибозо-5-фосфата и восстановлением двух молекул НАДФ + (2 НАДФ + → 2 НАДФН . Н + )
Таким образом, при окислении молекулы глюкозы образуется 2 НАДФН . Н + и рибозо-5-фосфат. В некоторых клетках катаболизм глюкозы на этом и заканчивается.
1)глюкозо-6-фосфатдегидрогеназа — главный ключевой фермент;
2) 6-фосфоглюконатдегидрогеназа.
Значение окислительного этапа:
1. Главный поставщик рибозо-5-фосфата для биосинтетических процессов:
— биосинтез мононуклеотидов (АМФ, ГМФ, УМФ, ЦМФ, ТМФ и др.);
— синтез нуклеиновых кислот (ДНК, РНК);
— синтез коферментов (НАД + , НАДФ + , ФАД, КоА-SН).
2. Основной источник НАДФН . Н + в клетках. ПФП на 50 % обеспечивает потребности клетки в НАДФН . Н + .
НАДФН . Н + в клетках используется:
1) в реакциях биосинтеза веществ как восстановитель:
— биосинтез холестерола, стероидных гормонов, желчных кислот;
— синтез заменимых аминокислот (НАДФН·Н + как кофермент глутаматдегидрогеназы в реакциях восстановительного аминирования α-кетоглутаровой кислоты);
2) в обезвреживании веществ: в реакциях гидроксилирования различных ксенобиотиков, лекарственных веществ, этанола и других веществ, которые осуществляются с участием микросомной цитР450-зависимой системы окисления;
3) как антиоксидант: используется на восстановление окисленного глутатиона. Глутатион — важного антиоксиданта клеток;
4) в фагоцитозе: генерирование активных форм кислорода. Фагоциты с использованием НАДФН . Н + генерируют супероксидные анион-радикалы, выполняющие основную роль в разрушении поглощённых бактериальных клеток. При недостаточной продукции НАДФН . Н + при нарушении ПФП отмечается хроническое течение инфекционных заболеваний.
Интенсивность протекания реакций ПФП зависит от потребности клеток в продуктах реакций и различается в разных тканях. Реакции окислительного этапа активно протекают в клетках печени, жировой ткани, эмбриональной ткани, в коре надпочечников, щитовидной железе, половых железах, лактирующей молочной железе, костном мозге, эритроцитах.
II. Неокислительный этап (этап межмолекулярных перегруппировок). На этом этапе происходят взаимопревращения сахаров (фосфотриоз, фосфотетроз, фосфопентоз, фосфогексоз, фосфогептулоз, фосфооктулоз), в результате которых регенерирует глюкозо-6-фосфат.
Два основных фермента катализируют превращения на неокислительном этапе:
1)транскетолазакатализирует перенос двухуглеродных фрагментов. В качестве кофермента использует тиаминпирофосфат;
2) трансальдолазакатализирует перенос трёхуглеродных фрагментов.
Варианты неокислительных превращений:
· классический или F-вариант (от англ. fat — жир) — осуществляется в клетках жировой ткани;
· октулозный или L-вариант (от англ. liver — печень) — осуществляется в клетках печени и других тканей.
Итак, на неокислительном этапе невостребованные в клетках пентозофосфаты в результате межмолекулярных перегруппировок превращаются в Гл-6-Ф, а также образуются Фр-6-Ф и 3-ФГА.
Все реакции неокислительного этапа обратимы.
На неокислительном этапе ПФП связан с гликолизом (посредством Гл-6-Ф, Фр-6-Ф и 3-ФГА), то есть возможно переключение этих процессов.
Значение неокислительного этапа:
1. Стабилизирует концентрацию фосфопентоз в клетке, то есть утилизирует лишние фосфопентозы. Благодаря связи с гликолизом лишние пентозы катаболизируют по гликолитическому пути, давая клеткам энергию.
2. Синтез фосфопентоз в клетке при торможении окислительного этапа благодаря обратимости реакций неокислительного превращения.
Регуляция пентозофосфатного пути, в основном, осуществляется на уровне дегидрогеназ. Инсулин индуцирует синтез глюкозо-6-фосфатдегидрогеназы, 6-фосфоглюконат-дегидрогеназы. Жирные кислоты — аллостерические ингибиторы глюкозо-6-фосфат-дегидрогеназы. Увеличение уровня НАДФН . Н + в клетке тормозит окисление глюкозы по ПФП.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом