
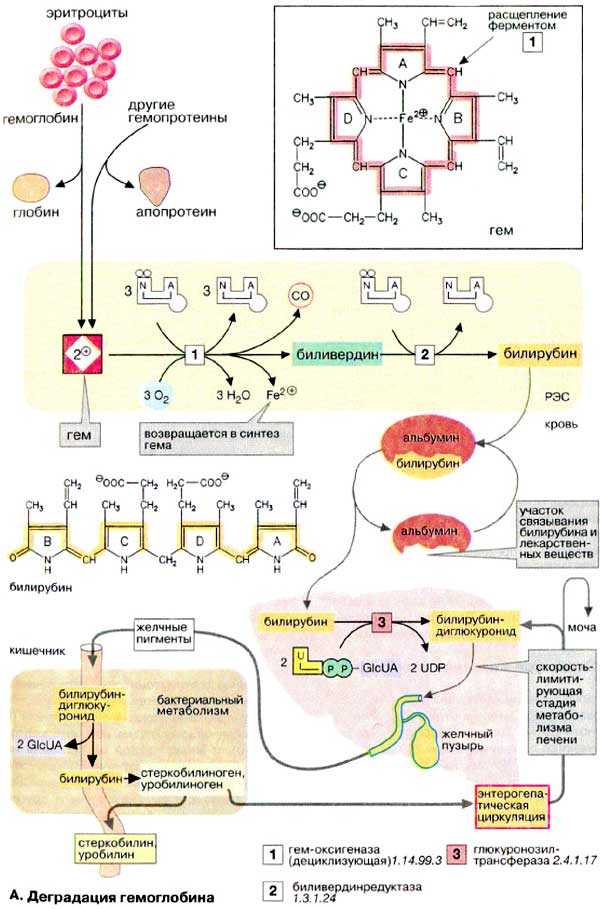
А. Деградация гемоглобина
В организме человека в течение 1 ч разрушается примерно 100-200 млн эритроцитов. Разрушение начинается в микросомальной фракции ретикуло-эндотелиальной системы [РЭС (RES)] клеток печени, селезенки и костного мозга. После отделения белковой части (глобина) красный гем расщепляется гем-оксигеназой с помощью кислорода и НАДФН на ионы Fe 2+ , СО (оксид углерода!) и зеленый биливердин . Далее железо утилизируется.
Затем биливердин восстанавливается биливердинредуктазой до оранжевого билирубина . Это изменение цвета легко можно наблюдать in vivo в виде синяков (гематомах). Интенсивный цвет гема и других порфиринов (см. рис. 195) является результатом сопряжения многочисленных двойных связей, которые образуют две резонансно стабилизированные (мезомерные) системы.
Для дальнейшего разрушения билирубин транспортируется кровью в печень. Так как он плохо растворим в плазме, транспорт осуществляется в комплексе с альбумином . В том же участке связывания альбумина сорбируются и лекарственные препараты. Паренхиматозные клетки печени забирают билирубин из крови.
После того как билирубин в печени дважды конъюгируется с активированной глюкуроновой кислотой (УДФ-GIcUA; см. рис. 113) (не показано), повышается его водорастворимость. Образование конъюгата катализируется УДФ-глюкуронозилтрансферазой — ферментом, находящимся в ЭР печени, а также в незначительных количествах в почках и слизистой кишечника. Глюкуроновая кислота присоединяется к пропионатным боковым цепям билирубина сложноэфирными связями. Образующийся диглюкуронид билирубина переносится в желчь путем активного транспорта против градиента концентрации. Этот транспорт является скорость-лимитирующей стадией метаболической трансформации билирубина в печени. Лекарственные препараты, такие, как, например фенобарбитал , могут индуцировать образование конъюгата и транспортный процесс.
В кишечнике конъюгат билирубина снова частично расщепляется бактериальной β-глюкуронидазой. Свободный билирубин постепенно восстанавливается до бесцветного уробилиногена и стеркобилиногена , которые далее окисляются кислородом воздуха до уробилина и стеркобилина. Эти конечные продукты метаболической трансформации желчных пигментов в кишечнике окрашены в цвета от оранжевого до желтого. Они выделяются по большей части с калом, а в меньшей степени резорбируются ( энтерогепатическая циркуляция; см. рис. 307). При интенсивном процессе разрушения гема в моче внезапно появляется уробилиноген, где он при окислении кислородом воздуха темнеет, превращаясь в уробилин .
Наряду с гемоглобином, по аналогичному пути разрушаются группы гема и у других гемсодержащих белков (миоглобина, цитохрома, каталазы, пероксидазы). Однако их вклад в образование желчных пигментов (250 мг в сутки) составляет лишь примерно 10-15%.
Гипербилирубинемия. Повышенный уровень билирубина ( >10 мг/л ) называется гипербилирубинемией. Билирубин диффундирует из крови в периферические ткани и окрашивает их в желтый цвет. Это особенно легко заметить на белой конъюктиве глаза, в таком случае говорят о желтухе. Ее причиной могут быть: повышенное образование билирубина из эритроцитов ( гемолитическая желтуха из-за наследственного дефекта фермента или отравления организма), нарушение выделения билирубина и продуктов его расщепления вследствие повреждений печени ( гепатоцеллюлярная желтуха из-за наследственного дефекта фермента или отравления организма) и застой желчи [ обтурационная (механическая) желтуха из-за желчных камней]. Неконъюгированный билирубин может даже проходить гематоэнцефалический барьер и приводить к поражению мозга ( ядерная желтуха ). Для точного диагноза причин гипербилирубинемии важен анализ билирубина в плазме. Конъюгированный («прямой») билирубин от неконъюгированного («непрямого») можно отличить с помощью цветной реакции.
Распад гема
Большая часть гемхромагенных пигментов в организме человека образуется при распаде гема. Главным источником гема является гемоглобин. В эритроцитах содержание гемоглобина составляет 80%, время жизни эритроцита 110-120 дней. В течение суток в организме взрослого человека массой 70 кг распадается около 6г гемоглобина. Распад эритроцитов происходит в клетках мононуклеарных фагоцитов. Наибольшее число таких клеток в селезенке, печени, костном мозге.
Первая реакция распада гемоглобина – это гидролиз его белковой части.
Катаболизм гема, освобождение от белковой части происходит в микросомальной фракции при участии сложной гем-оксигеназной ферментативной системы, требующей наличие НАДФН2 и кислорода. При поступлении гема в гемаксиназную систему микросом ионы железа окисляются в ферри форму, т.е. гем превращается в гемин.
гем (Fe 2+ ) (над: оксигеназа) гемин(Fe 3+ ) (это ферри форма) (над: редуктаза; под: НАДФH2НАДФ) гемин (Fe 2+ ) (ферро форма) (над: редуктаза, +О2; под: НАДФН2НАДФ)оксигемин (Fe 3+ ) (над: редуктаза, +О2; под: -СО, -Fe 3+ )биливердин ( над: НАДФН2НАДФ) билирубин (над: +альбумин) билирубин-альбумин (неконъюгированный билирубин — НБ).
Как видно из схемы, в процессе реакций катаболизма гема, гемин восстанавливается с помощью редуктаз (НАДФН2) в ферро-форму (Fe 2+ ).
При участии НАДФН2, кислород проходит последующее превращение с раскрытием тетрапирольного кольца гема, с выделением окиси углерода (СО) и освобождением ферри-иона (Fe 3+ ).
Образуется ациклическое соединение биливердин, у которого редуктаза восстанавливает метиловый мостик между 3 и 4 пирролами, образуется желтый, токсический для клеток, пигмент – билирубин.
За сутки у взрослого человека массой 70 кг образуется 250-350 мг билирубина. Химические превращения гема в билирубин в клетках МФ можно наблюдать внутри организма в гематоме, где пурпурный цвет, обусловленный гемом, медленно переходит в желтый цвет, обусловленный билирубином.
Образованный в клетках билирубин является токсическим веществом, удаляется из них и поступает в кровь, взаимодействуя с транспортными белками — альбуминами. Образованное комплексное соединение билирубин-альбумин называется неконъюгированный билирубин — НБ.
3. адсорбирован на альбумине;
4. не проходит через почечный эпителий;
5. не дает прямой реакции с диазо-реактивом Эрлиха.
НБ с помощью альбумина поступает для детоксикации в печень, где в гепатоцитах, в реакции конъюгации с глюкуроновой кислотой, при участии ТФ образуются:
Эти билирубины носят название конъюгированного билирубина – КБ.
4. легко проникает через почечный барьер;
5. дает прямую реакцию с диазо-реактивом Эрлиха.
Этот билирубин (КБ) может проникать в кровяные капилляры. В плазме крови на его долю приходиться 25%, от общего билирубина, который в норме составляет 8-20 мкмоль/л.
Далее из печени КБ в составе желчи поступает в клетки, где под влиянием ферментов микрофлоры он гидролизуется. Отщепившаяся глюкуроновая кислота всасывается в слизистую кишечника и через воротную вену вновь поступает в печень, где может использоваться для детоксикации.
Читайте также: Шаблон для мяча из ткани
Билирубин под влиянием ферментов кишечной микрофлоры многократно восстанавливается, превращается в мезобилиноген, часть которого может всасываться слизистой и через систему воротной вены поступать в печень, где разрушается до моно-, дипирролов, которые из организма удаляются в составе желчи с каловыми массами.
Незначительная часть мезобилиногена с током крови поступает в почки, где превращается в другой пигмент – уробилиноген, который, окисляясь, образует пигмент мочи – уробилин.
В моче здорового взрослого человека массой 70 кг уробилин присутствует в следовых количествах.
За сутки (суточный диурез в норме равен 1,2-1,5 л) выделяет 1-4 мг уробилина.
Большая часть мезобилиногена в толстом отделе кишечника под влиянием ферментов кишечной микрофлоры, восстанавливаясь, превращается в стеркобилиноген – основной пигмент кала, который, окисляясь, превращается в стеркобилин. За сутки с калом выводиться примерно 280-300 мг стеркобилина.
29. Катаболизм гема в организме.
За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов. Эритроциты живут 90-120 дней, после чего лизируются в кровеносном русле или в селезенке.При разрушении эритроцитов в кровяном русле высвобождаемый гемоглобин образует комплекс с белком-переносчиком гаптоглобином (фракция α2-глобулинов крови) и переносится в клетки ретикуло-эндотелиальной системы (РЭС) селезенки (главным образом), печени и костного мозга.
В клетках РЭС гем в составе гемоглобина окисляется молекулярным кислородом. В реакциях последовательно происходит разрыв метинового мостика между 1-м и 2-м пиррольными кольцами гема с их восстановлением, отщеплением железа и белковой части и образованием оранжевого пигмента билирубина.
Билирубин – токсичное, жирорастворимое вещество, способное нарушать окислительное фосфорилирование в клетках. Особенно чувствительны к нему клетки нервной ткани.
Из клеток ретикуло-эндотелиальной системы билирубин попадает в кровь. Здесь он находится в комплексе с альбумином плазмы, в гораздо меньшем количестве – в комплексах с металлами, аминокислотами, пептидами и другими малыми молекулами. Образование таких комплексов не позволяет выделяться билирубину с мочой. Билирубин в комплексе с альбумином называется свободный (неконъюгированный) или непрямой билирубин. Из сосудистого русла в гепатоциты билирубин попадает с помощью белков-переносчиков. Далее при участии белка лигандина он транспортируется в ЭПР, где протекает реакция связывания билирубина с УДФ-глюкуроновой кислотой, при этом образуются моно- и диглюкурониды. Кроме глюкуроновой кислоты, в реакцию могут вступать сульфаты, фосфаты, глюкозиды. Билирубин-глюкуронид получил название связанный (конъюгированный) или прямой билирубин.После образования билирубин-глюкурониды АТФ-зависимым переносчиком секретируются в желчные протоки и далее в кишечник, где при участии бактериальной β-глюкуронидазы превращаются в свободный билирубин.Одновременно некоторое количество билирубин-глюкуронидов может попадать (особенно у взрослых) из желчи в кровь по межклеточным щелям.Таким образом, в плазме крови в норме одновременно существуют две формы билирубина: свободный (непрямой), попадающий сюда из клеток РЭС (около 80% всего количества), и связанный (прямой), попадающий из желчных протоков (до 20%).
В кишечнике билирубин подвергается восстановлению под действием микрофлоры до мезобилирубина и мезобилиногена (уробилиногена). Часть последних всасывается и с током крови вновь попадает в печень, где окисляется до ди- и трипирролов. При этом в здоровом организме в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами. Оставшаяся в кишечнике часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена и выделяется из организма, окрашивая кал. Незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда в почки и выделяется с мочой. На воздухе стеркобилиноген и уробилиноген превращаются, соответственно, в стеркобилин и уробилин.
30. Обмен гемоглобина, его синтез и распад, образование желчных пигментов, их нормальное содержание, диагностическое значение определения желчных пигментов в крови и моче. Гемоглобин входит в состав группы белков гемопротеины, которые сами являются подвидом хромопротеинов и подразделяются на неферментативные белки (гемоглобин, миоглобин) и ферменты (цитохромы, каталаза, пероксидаза). Небелковой частью их является гем структура, включающаявсебяпорфириновоекольцо (состоящее из 4 пиррольных колец) и иона Fe2+. Железо связывается с порфириновым кольцом двумя координационными и двумя ковалентными связями.
Строение гемоглобина. Гемоглобин представляет собой белок, включающий 4 гемсодержащие белковые субъединицы. Между собой протомеры соединяются гидрофобными, ионными, водородными связями по принципу комплементарности. При этом они взаимодействуют не произвольно, а определенным участком — контактной поверхностью. Этот процесс высокоспецифичен, контакт происходит одновременно в десятках точек по принципу комплементарности. Взаимодействие осуществляют разноименно заряженные группы, гидрофобные участки, неровности на поверхности белка. Белковые субъединицы в нормальном гемоглобине могут быть представлены различными типами полипептидных цепей: α, β, γ, δ, ε, ξ (соответственно, греч. — альфа, бета, гамма, дельта, эпсилон, кси). В состав молекулы гемоглобина входят по две цепи двух разных типов. Гем состоит из иона двухвалентного железа и порфирина. В основе структуры порфиринов находится порфин. Порфин представляет собой четыре пиррольных кольца, связанных между собой метеновыми мостикамиГем соединяется с белковой субъединицей, во-первых, через остаток гистидина координационной связью железа, во-вторых, через гидрофобные связи пиррольных колец и гидрофобных аминокислот. Гем располагается как бы «в кармане» своей цепи и формируется гемсодержащий протомер.
Существует несколько нормальных вариантов гемоглобина:
HbР примитивныйгемоглобин, содержит 2ξ- и 2ε-цепи, встречаетсяв эмбрионе между 7-12 неделями жизни,
HbF фетальныйгемоглобин, содержит 2α- и 2γ-цепи, появляетсячерез 12 недельвнутриутробногоразвитияиявляетсяосновнымпосле 3 месяцев,
HbA гемоглобинвзрослых, долясоставляет 98%, содержит 2α- и 2β-цепи, уплода появляется через 3 месяца жизни и к рождению составляет 80% всего гемоглобина,
HbA2 гемоглобинвзрослых, долясоставляет 2%, содержит 2α- и 2δ-цепи,
HbO2 оксигемоглобин, образуетсяприсвязываниикислородавлегких, влегочныхвенахего 94-98% отвсего количества гемоглобина,
HbCO2 карбогемоглобин, образуетсяприсвязыванииуглекислогогазавтканях, ввенознойкровисоставляет 15-20% отвсегоколичествагемоглобина.
Патологические формы гемоглобина
MetHb метгемоглобин, формагемоглобина, включающаятрехвалентныйионжелезавместодвухвалентного. Такаяформаобычнообразуетсяспонтанно, вэтомслучаеферментативныхмощностейклеткихватаетнаеговосстановление. Прииспользованиисульфаниламидов, употреблении нитрита натрия и нитратов пищевых продуктов, при недостаточности аскорбиновой кислоты ускоряется переход Fe2+ в Fe3+. Образующийся metHb не способен связывать кислород и возникает гипоксия тканей. Для восстановления ионов железа в клинике используют аскорбиновую кислоту и метиленовую синь.
Читайте также: Таблица всех тканей растений
Hb-CO карбоксигемоглобин, образуетсяприналичииСО (угарныйгаз) вовдыхаемомвоздухе. Онпостоянноприсутствуетвкровивмалыхконцентрациях, ноегодоляможетколебатьсяотусловийиобразажизни. Угарныйгаз является активным ингибитором гем-содержащих ферментов, в частности, цитохромоксидазы 4-го комплекса дыхательной цепи.
HbA1С гликозилированныйгемоглобин. Концентрацияегонарастаетприхроническойгипергликемиииявляетсяхорошимскрининговымпоказателем уровня глюкозы крови за длительный период времени.
Биосинтез гема. Гем является небелковой частью многих гемопротеинов:
гемоглобин (до 85% общего количества гема организма), локализованный в эритроцитах и клетках костного мозга,миоглобин скелетных мышц и миокарда (17%), цитохромы дыхательной цепи и P450, цитохромоксидаза, гомогентизатоксидаза, пероксидаза, миелопероксидаза, каталаза, тиреопероксидаза и т.д. менее 1%. Синтезгемавосновномидетвпредшественникахэритроцитов, клеткахпечени, почек, слизистой кишечника, и в остальных тканях. Первая реакция синтеза с участием δ-аминолевулинат-синтазы происходит в митохондриях. Следующая реакция при участии аминолевулинатдегидратазы (порфобилиноген-синтазы) протекает в цитозоле.Гем синтезируется во всех тканях, но с наибольшей скоростью в костном мозге и печени. В костном мозге гем необходим для синтеза гемоглобина в ретикулоцитах, в гепатоцитах — для образования цитохрома Р450.
Первая реакция синтеза гема — образование 5-аминолевулиновой кислоты из глицина и сук-цинил-КоА идёт в матриксе митохондрий, где в ЦТК образуется один из субстратов этой реакции — сукцинил-КоА. Эту реакцию катализирует пиридоксальзависимый фермент аминолевулинатсинтаза. Из митохондрий 5-аминолевулиновая кислота поступает в цитоплазму. В цитоплазме проходят промежуточные этапы синтеза гема: соединение 2 молекул 5-аминолевулиновой кислоты молекулу порфобилиногена, дезаминирование порфобилиногена с образованием гидроксиметилбилана, ферментативное превращение гидроксиметилбилана в молекулу уропор-фобилиногена III, декарбоксилирование последнего с образованием копропорфириногена III. Гидроксиметилбилан может также нефермента-тивно превращаться в уропорфириноген I, который декарбоксилируется в копропорфирино-ген I. Из цитоплазмы копропорфириноген III опять поступает в митохондрии, где проходят заключительные реакции синтеза гема. В результате двух последовательных окислительных реакций копропорфириноген III превращается в протопорфириноген IX, а протопорфириноген IX — в Протопорфирин IX. Фермент феррохела-таза, присоединяя к протопорфирину IX двухвалентное лентное железо, превращает его в гем. Источником железа для синтеза гема служит депонирующий железо белок ферритин. Синтезированный гем, соединяясь с α и β-полипепептидными цепями глобина, образует гемоглобин. Гем регулирует синтез глобина: при снижении скорости синтеза гема синтез глобина в ретикулоцитах тормозится.
Распад. За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов. Эритроциты живут 90-120 дней, после чего лизируются в кровеносном русле или в селезенке. При разрушении эритроцитов в кровяном русле высвобождаемый гемоглобин образует комплекс с белком-переносчиком гаптоглобином (фракция α2-глобулинов крови) и переносится в клетки ретикуло-эндотелиальной системы (РЭС) селезенки (гл образом), печени и костного мозга.Первая реакция катаболизма гема происходит при участии NADPH-зависимого ферментативного комплекса гемоксигеназы. Ферментная сисгема локализована в мембране ЭР, в области электронтранспортных цепей микросомального окисления. Фермент катализирует расщепление связи между двумя пиррольными кольцами, содержащих винильные остатки, — таким образом, раскрывается структура кольца. В ходе реакции образуются линейный тетрапир-рол — биливердин (пигмент жёлтого цвета) и монооксид углерода (СО), который получается из углерода метениловой группы. Гем индуцирует транскрипцию гена гемоксигеназы, абсолютно специфичной по отношению к тему.Ионы железа, освободившиеся при распаде гема, могут быть использованы для синтеза новых молекул гемоглобина или для синтеза других железосодержащих белков. Биливердин восстанавливается до билирубина NADPH-зависимым ферментом биливердинредуктазой. Билирубин образуется не только при распаде гемоглобина, не также при катаболизме других гемсодержащю белков, таких как цитохромы и миоглобин. При распаде 1 г гемоглобина образуется 35 мг билирубина, а в сутки у взрослого человека — примерно 250-350 мг билирубина. Дальнейший метаболизм билирубина происходит в печени. В клетках РЭС гем в составе гемоглобина окисляется молекулярным кислородом. В реакциях последовательно происходит разрыв метинового мостика между 1-м и 2-м пиррольными кольцами гема с их восстановлением, отщеплением железа и белковой части и образованием оранжевого пигмента билирубина. Билирубин токсичное, жирорастворимоевещество, способноенарушатьокислительноефосфорилированиевклетках. Особенночувствительныкнемуклеткинервнойткани. Изклетокретикуло-эндотелиальнойсистемыбилирубинпопадаетвкровь. Здесьон находится в комплексе с альбумином плазмы, в гораздо меньшем количестве вкомплексахсметаллами, аминокислотами, пептидамиидругимималымимолекулами. Образованиетакихкомплексовнепозволяетвыделятьсябилирубинусмочой. Билирубинвкомплексесальбумином называется свободный (неконъюгированный) или непрямой билирубин. Из сосудистого русла в гепатоциты билирубин попадает с помощью белка-переносчика (лигандина). В клетке протекает реакция связывания билирубина с УДФ-глюкуроновой кислотой, при этом образуются моно- и диглюкурониды. Кроме глюкуроновой кислоты, в реакцию могут вступать сульфаты, фосфаты, глюкозиды. Билирубин-глюкуронид получил название связанный (конъюгированный) или прямой билирубин. После образования билирубин-глюкурониды АТФ-зависимым переносчиком секретируются в желчные протоки и далее в кишечник, где при участии бактериальной β-глюкуронидазы превращаются в свободный билирубин. Одновременно некоторое количество билирубин-глюкуронидов может попадать (особенно у взрослых) из желчи в кровь по межклеточным щелям. Таким образом, в крови в норме одновременно существуют две формы билирубина: свободный, попадающий сюда из клеток РЭС (около 80% всего количества), и связанный, попадающий из желчных протоков (до 20%). Превращение в кишечнике. В кишечнике билирубин подвергается восстановлению под действием микрофлоры до мезобилирубина и мезобилиногена (уробилиногена). Часть последних всасывается и с током крови вновь попадает в печень, где окисляется до ди- и трипирролов.
Жёлчные (или же́лчные) пигме́нты, также билины — биологические пигменты, линейные тетрапирролы[1], формально являющиеся производными билана (билиногена) с окисленными терминальными пиррольными ядрами, образующиеся при катаболизме гема. Впервые выделены из жёлчи, которой придают характерную окраску, откуда и получили своё название; цвет различных жёлчных пигментов — от жёлто-оранжевого до сине-зелёного. Образуются во многих организмах как продукт метаболизма некоторых порфиринов. Билин (также называемый билихром) был назван как желчный пигмент млекопитающих, но его также можно обнаружить в низших позвоночных, беспозвоночных, а также в красных водорослях, зелёных растениях и цианобактериях. Цвет билинов может варьировать от красного, оранжевого, жёлтого и коричневого до голубого и зелёного.Говоря химическим языком, билины это линейная структура из четырёх пиррольных колец (тетрапирролы). В человеческом метаболизме они представлены билирубином-продуктом разрушения гема. Гидроксиметилбилан это одно из популярных анаболических средств, получаемое реакцией биосинтеза порфобилиногена (ПБГ) и уропорфобилиногена I (реакция широко известена как порфобилиногенная деаминаза).
При диагностике желтух надо иметь в виду, что на практике редко отмечают желтуху какого-либо одного типа в «чистом» виде. Чаще встречается сочетание того или иного типа. Так, при выраженной гемолитической желтухе, сопровождающейся повышением концентрации непрямого билирубина, неизбежно страдают различные органы, в том числе и печень, что может вносить элементы паренхиматозной желтухи, т.е. повышение в крови и моче прямого билирубина. В свою очередь, паренхиматозная желтуха, как правило, включает в себя элементы механической. При подпечёночной (механической) желтухе, например при раке головки поджелудочной железы, неизбежен повышенный гемолиз как следствие раковой интоксикации и, как следствие, повышение в крови как прямого, так и непрямого билирубина. Итак, гипербилирубинемия может быть следствием избытка как связанного, так и свободного билирубина. Измерение их концентраций по отдельности необходимо при постановке диагноза желтухи. Если концентрация билирубина в плазме – + H +
Читайте также: Снижение содержания кислорода в тканях это
Остальные бикарбонаты выходят в плазму крови в обмен на ионы хлора (гипохлоремический сдвиг). В легочных капиллярах имеется относительно низкая концентрация углекислого газа в альвеолярном воздухе, происходит высокоэффективная диффузия СО2 из плазмы через альвеолярные мембраны и его удаление с выдыхаемым воздухом . Уменьшение концентрации СО2 в плазме стимулирует его образование в карбоангидразной реакции внутри эритроцита и снижает здесь концентрацию иона HCO3 – .
Одновременно высокая концентрация кислорода вытесняет СО2 из комплекса с гемоглобином с образованием оксигемоглобина – более сильной кислоты, чем угольная кислота . Диссоциирующие от оксигемоглобина ионы Н+ нейтрализуют поступающий извне ион HCO3 – с образованием угольной кислоты. После карбоангидразной реакции образуется СО2, который выводится наружу.
(36-37) Что такое гипоксия, ее виды, причины возникновения?
Гипоксия (кислородное голодание) — представляет собой патологической состояние, характеризующееся дефицитом кислорода в организме, которое возникает вследствие его недостаточного поступления извне или на фоне нарушения процесса утилизации на клеточном уровне.
Дыхательная (респираторная) гипоксия развивается при заболеваниях органов дыхания (например, бронхиты, легочная гипертензия, любые патологии легких и т.д.), когда затрудняется проникновение кислорода из воздуха в кровь. На фоне респираторной гипоксии могут развиться осложнения, такие, как дыхательная недостаточность, отек мозга и газовый ацидоз.
Циркуляторная (сердечно-сосудистая) гипоксия развивается на фоне различных расстройств кровообращения (например, снижения тонуса сосудов, уменьшения общего объема крови после кровопотери или обезвоживания, повышения вязкости крови, усиления свертываемости, централизации кровообращения, венозного застоя и т.д.). При циркуляторной гипоксии через легкие в кровь поступает нормальное количество кислорода, но из-за нарушения кровообращения он с опозданием доставляется к органам и тканям, вследствие чего в последних возникает кислородное голодание. По механизму развития циркуляторная гипоксия бывает ишемической и застойной.
Гемическая (кровяная) гипоксия развивается при нарушении качественных характеристик или уменьшении количества гемоглобина крови. Гемическая гипоксия подразделяется на две формы – анемическую и обусловленную изменениями качества гемоглобина. Анемическая гемическая гипоксия обусловлена снижением количества гемоглобина в крови, то есть анемией любого происхождения. А гипоксия, обусловленная изменением качества гемоглобина, связана с отравлением различными ядовитыми веществами, которые приводят к образованию форм гемоглобина, не способных переносить кислород (метгемоглобина или карбоксигемоглобина).
Тканевая (гистотоксическая) гипоксия развивается на фоне нарушения способности клеток органов поглощать кислород. Причиной тканевой гипоксии является сниженная активность или дефицит ферментов дыхательной цепи митохондрий, которые переводят кислород в формы, в которых он используется клетками для осуществления всех процессов жизнедеятельности.
(39) Роль постоянства КОС в нормальном функционировании организма. Нормальные показатели в крови. Охарактеризуйте буферные системы крови. Какие из них эритроцитарные, какие плазменные?
Наиболее эффективно поддерживают КОС легкие и почки, также задействованы печень, пищеварительный канал и костная ткань.
Кислотно-основное равновесие представляет собой активность физиологических и физико-химических процессов, составляющих функционально единую систему стабилизации концентрации ионов Н+. Нормальные величины концентрации ионов Н+ около 40 нмоль/л, что в 106 раз меньше, чем концентрация многих других веществ. Совместимые с жизнью колебания концентрации ионов Н+ располагаются в пределах 16-160 нмоль/л.
Так как реакции обмена веществ часто связаны с окислением и восстановлением молекул, то в этих реакциях обязательно принимают участие соединения, выступающие в качестве акцептора или донора ионов водорода. Роль других соединений – обеспечить неизменность концентрации ионов водорода при жизнедеятельности.
рН внутренних сред организма (крови, лимфы, ликвора, желудочного сока, мочи) оказывает воздействие на жизнедеятельность клеток, тканей, органов и организма в целом.
Значение рН внутренних сред характеризуется значительным постоянством и устойчивостью.
рН – это отрицательный десятичный логарифм концентрации ионов водорода.
(41) Бикарбонатная буферная система: состав, механизм действия, значение. Ацидозы и алкалозы: виды, причины.
Бикарбонатная буферная система является самой регулируемой системой крови. На ее долю приходится около 10% всей буферной емкости крови. Состоит из угольной кислоты Н2СО3 и бикарбонат-иона НСО3. При нормальном значении рН крови соотношение Н2СО3 к НСО3 – 1/20.
Бикарбонатная буферная система функционирует как эффективный регулятор в области рН 7,4.

Смещение КОС крови в сторону повышения концентрации ионов водорода (снижение рН до 7,0) и уменьшения резервной щелочности ― ацидоз. Смещение КОС крови в сторону понижения концентрации ионов водорода (повышение рН до 7,8) и увеличения резервной щелочности крови ―алкалоз.
Нарушения КОС: 1) Алкалоз (метаболический, респираторный)
2) Ацидоз (метаболический респираторный)
1. Повышение содержания кислот в крови
•Кетоацидоз (при сахарном диабете, недостатке углеводов в пище при достаточном потреблении белков и жиров, при отравлении алкоголем)
•Лактоацидоз (при сепсисе, кровотечении, отеке легких, сердечной недостаточности, шоке, при остром панкреатите, сахарном диабете, лейкемии, хроническом алкоголизме).
•С кишечным, панкреатическим и билиарным секретами при диареях и фистулах кишечника и желчного пузыря.
3. Недостаточное выведение ионов Н+ почками
•хронической почечной недостаточности или поражение канальцев.
Причинами являются нарушение вентиляции легких, сопровождающиеся гиповентиляцией:
•Повреждения или заболевания легких (пневмония, фиброз, отек легких)
•Повреждения или заболевания дыхательных мышц (нехватка калия, боли после операции, травмы, накопление жировых отложений)
•Угнетение дыхательного центра (опиаты, барбитураты), неправильный режим ИВЛ

Бронхиальная астма, эмфизема, бронхит.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом