
В каких тканях работает глицерофосфатный челночный механизм
Образующийся в цитоплазме NADH не может самостоятельно проникать через внутреннюю митохондриальную мембрану (ВММ) в матрикс (внутреннее пространство митохондрии), где он должен утилизироваться в дыхательной цепи, поставляя в нее электроны. Эту функцию выполняет целый ряд челночных механизмов, из которых два заслуживают наибольшего внимания.
Дигидроксиацетон -3- фосфат взаимодействует с цитоплазматическим NADH с образованием глицерин -3- фосфата:
СН 2 (ОН)-С(=О)-СН-ОРО 3 Н 2 + NADH + H+ = СН 2 (ОН)-СH(ОH)-СН-ОРО 3 Н 2 + NAD+
Глицерин -3- фосфат передает электроны на FMN, локализованный во внутримитохондриальной мембране, по схеме:
FMN + СН 2 (ОН)-СH(ОH)-СН-ОРО 3 Н 2 = СН 2 (ОН)-С(=О)-СН-ОРО 3 Н 2 + FMN.H2
Щавелевоуксусная кислота (ЩУК) взаимодействует с цитоплазматическим NADH с образованием малата (яблочной кислоты) и NAD + . Малат проникает через мембрану в матрикс и окисляется до оксалоацетата (ЩУК) митозольным NAD + , в результате чего образуется внутримитохондриальный NADH, который и поступает в дыхательную цепь.
Далее, ЩУК аминируется от глутамината до аспартата, который совершает переход через мембрану в цитоплазму, где обратно переаминируется с aльфа-кетоглутаратом до образования ЩУК. Альфа-кетоглутаровая кислота пересекает мембрану самостоятельно, без помощи каких-либо переносчиков. В результате всего происходит перенос двух электронов из цитозоля в митозоль по схеме:
NADHcytos + NAD + mytos = NADHmytos + NAD + cytos
Из перечисленных соединений аспартат-малатного челнока только оксалоацетат и NADH не могут самостоятельно пересекать ВММ (точнее, не имеют встроенных в эту мембрану переносчиков). Для обеспечения функционирования челнока и требуются реакции переаминирования.
Реакция переаминирования катализируется ГЛУТАМАТ-ОКСАЛОАЦЕТАТ-ТРАНСАМИНАЗОЙ.
Разница между двумя механизмами переноса восстановительных эквивалентов заключается в том, что аспартат-малатный челнок поставляет в дыхательную цепь молекулы NADH, фигурирующие в начальных ее этапах. При переносе по дыхательной цепи пары электронов, полученных из молекулы NADH, синтезируется в итоге 3 молекулы АТФ. Глицерофосфатный челнок поставляет в цепь молекулы FAD.H2, в результате чего из одной цитоплазматической NADH синтезируется только 2 молекулы АТФ (FAD.H2 располагается на более низких «энергетических ступенях» дыхательной цепи).
Челночные механизмы переносят в митозоль восстановительные эквиваленты образованные не только в реакциях гликолиза, но также и из других окислительно-восстановительных реакций, например:
С 2 Н 5 ОН + NAD + = CH3CHO + NADH + H +
образующиеся в результате окисления этанола в цитозоле клеток печени NADH должны быть перенесены в митозоль (матрикс митохондрий) для утилизации в дыхательной цепи. Это осуществляется посредством одного из челночных механизмов. Дальнейшее окисление ацетальдегида протекает в матриксе ацетальдегиддегидрогеназой. Ацетальдегид достигает матрикса без помощи переносчиков.
При утилизации печенью различных чужеродных для организма лекарственных метаболитов также используются челночные механизмы. При этом происходят следующие процессы. УДФ-глюкоза окисляется в цитоплазме до УДФ-глюкуроновой кислоты:
УДФ-глюкоза + NAD+ = УДФ-глюкуроновая кислота + NADH + H+
УДФ-глюкуронат связывает и выводит через почки метаболиты по схеме:
УДФ-глюкуронат(СООН) + НО-R = УДФ + глюкуронат-метаболит
Реакция катализируется трансферазами. В результате переноса на метаболит остатка углевода его растворимость в воде резко повышается. Растворимость глюкуронатов в воде, тем более, повышена вследствие наличия слабодиссоциирующей карбоксильной группы СООН. Хорошая растворимость в воде является необходимым условием быстрого выведения метаболита через почки.
Однако, производительность челноков не беспредельна и совместное принятие лекарственных средств и алкоголя может приводить к их перегрузке, и как следствие, к пролонгации действия лекарств, в результате замедления механизмов выведения. Кроме того, алкоголь в ряде случаев приводит к усилению действия лекарств в результате синергического эффекта. Терапевтические дозы препаратов могут оказаться опасными для жизни в присутствии алкоголя.
Лица, страдающие хроническим алкоголизмом, обладают, будучи в трезвом состоянии, повышенной способностью метаболировать чужеродные организму соединения, в результате развития кросс-толерантности по причине частого употребления алкоголя и индуцирования большого количества утилизирующих ферментов. В результате развивается следующий сценарий. Он/Она имеет проблемы со сном. Обычная терапевтическая доза барбитуратов не приносит желаемого эффекта. В прострации выпивается одна за другой несколько пилюль и все запивается алкоголем. Наступает глубокий сон, но в результате сильного угнетения ЦНС останавливается дыхание и наступает смерть, поскольку в комбинации с алкоголем устойчивость к действию барбитуратов утрачивается.
Читайте также: Как сшить глобус из ткани
НАДН гликолиза могут доставляться в митохондрии
Молекулы НАДН, образованные в шестой реакции гликолиза, в зависимости от наличия кислорода имеют, как минимум, два пути своего дальнейшего превращения:
- либо остаться в цитозоле и вступить в одиннадцатую реакцию гликолиза ( анаэробные условия),
- либо проникнуть в митохондрию и окислиться в дыхательной цепи ( аэробные условия).
Челночные системы
Так как сама молекула НАДН через мембрану не проходит, то существуют специальные системы, принимающие атомы водорода от НАДН в цитоплазме и отдающие их в матриксе митохондрий. Эти системы получили название челночные системы .
Определены две основные челночные системы – глицеролфосфатная и малат-аспартатная.
Глицеролфосфатный челночный механизм
Главными ферментами глицеролфосфатного челнока являются изоферменты глицерол-3-фосфат-дегидрогеназы – цитоплазматический и митохондриальный. Они отличаются своими коферментами: у цитоплазматической формы – НАД, у митохондриальной – ФАД.
В цитозоле метаболиты гликолиза – диоксиацетонфосфат и НАДН образуют глицерол-3-фосфат , поступающий в митохондрии. Там он окисляется с образованием ФАДН2. Далее ФАДН2 направляется в дыхательную цепь и используется для получения энергии. Таким образом, в результате действий челнока цитозольный НАДН+H + как бы «превращается» в митохондриальный ФАДН2.
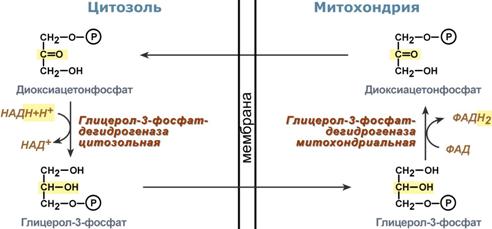
Схема работы глицерол-фосфатной челночной системы
Этот челнок активен в печени, в белых скелетных мышцах и в бурой жировой ткани. Однако в гепатоците в состоянии покоя и после еды часть глицерол-3 фосфата в митохондрию не пойдет, а будет использоваться в цитозоле для синтеза фосфолипидов и триацилглицеролов.
Малат-аспартатный челночный механизм
Главными ферментами этого челнока являются изоферменты малатдегидрогеназы – цитоплазматический и митохондриальный. Он является распространенным по всем тканям.
Этот механизм более сложен: постоянно идущие в цитоплазме при участии фермента аспартатаминотрансферазы ( АСТ ) реакции трансаминирования аспарагиновой кислоты с α-кетоглутаратом поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназы и за счет «гликолитического» НАДН восстанавливается до яблочной кислоты (малата).
Последняя антипортом с α-кетоглутаратом проникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он при помощи аспартатаминотрансферазы трансаминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль.
Таким образом, атомы водорода от цитозольного НАДН перемещаются в состав митохондриального НАДН.
Челночные механизмы для НАДH
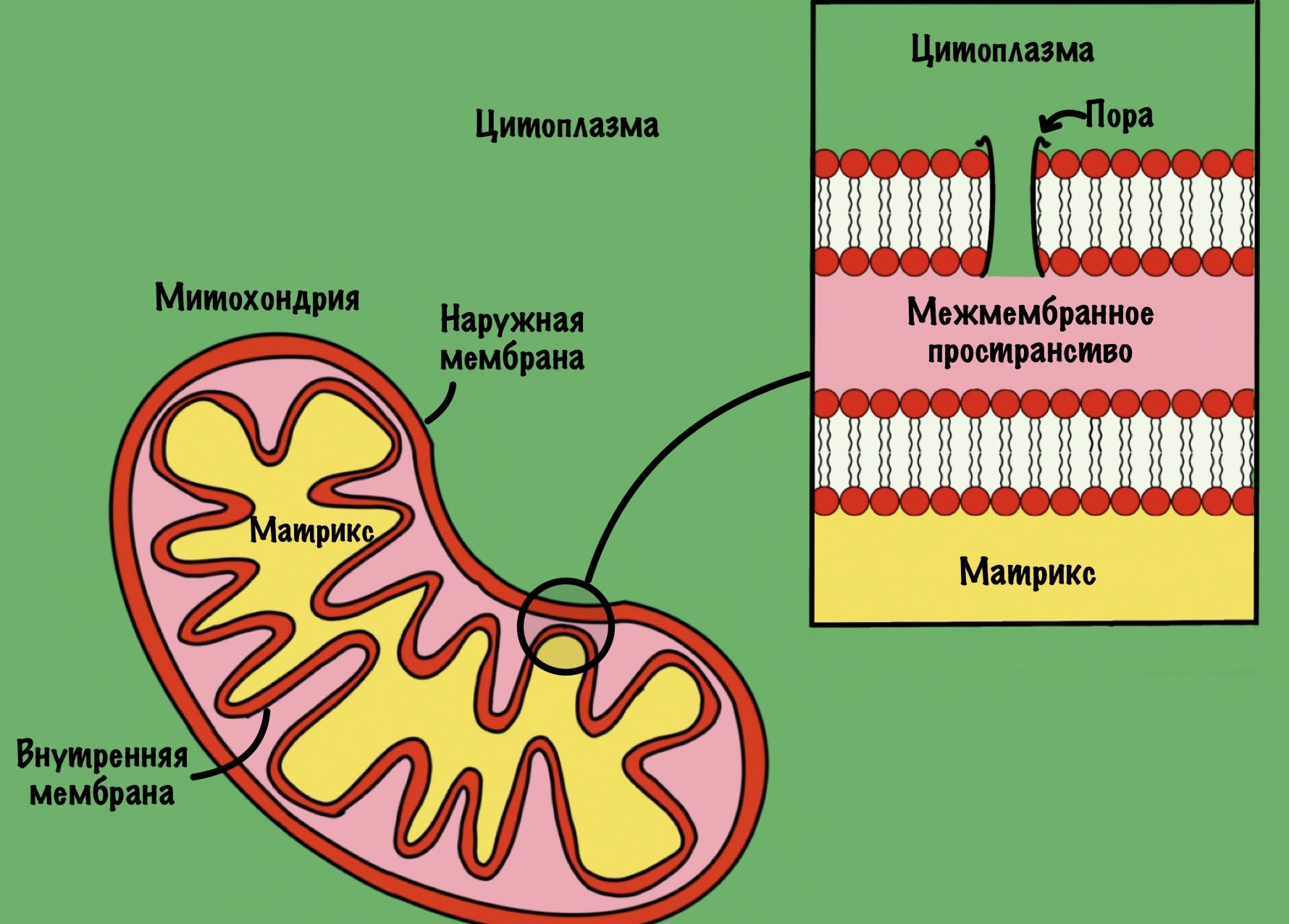
Челночные механизмы нужны для переноса электронов и протона водорода с НАДH в матрикс митохондрий. Почему эта молекула так важна, что для неё существуют отдельные механизмы?
НАДH — это восстановленный кофермент. Он образовался в шестой реакции гликолиза, которую катализирует глицеральдегидфосфатдегидрогеназа. Для этой реакции в активном центре фермента должен находиться кофермент — НАД + . Кофермент принимает на себя два электрона и протон от глицеральдегид-3-фосфата. Так образуется восстановленный НАДH.
Шестая реакция гликолиза не пойдёт, если в активном центре фермента не будет НАД + . А это значит, что реакции гликолиза остановятся. Так, если следующая глюкоза попадает в гликолиз, то дальше пятой реакции дело не пойдёт. Клетка не получит то, ради чего всё затевалось: две молекулы АТФ, две молекулы пирувата и две молекулы НАДH. Обидно терять всё это богатство из-за одного кофермента.
Тут нужно остановиться и вспомнить, что гликолиз идёт в цитоплазме клетки. Там НАДH можно окислить с помощью анаэробного гликолиза. Но клетку такое интересует не часто. Из-за анаэробного гликолиза она теряет две молекулы пирувата, которые пойдут на синтез лактата. Эту реакцию катализирует лактатдегидрогеназа. С помощью неё НАДH окисляется до НАД + . Эта реакция очень полезна в мышцах, когда им не хватает кислорода, и эритроцитах. НО остаётся огромный минус — клетка не получит пируват. Он не отправится в цикл трикарбоновых кислот и дыхательную цепь, а значит куча энергии пропадёт даром.
Читайте также: Зона сжатия костной ткани
Нужен лучший способ окислить НАДH до НАД + , при котором мы не пожертвуем двумя молекулами пирувата. Для этого есть митохондрия, которая отделена двумя биологическими мембранами от цитоплазмы. Там основная энергетическая топка нашей клетки. При наличии кислорода восстановленные коферменты (НАДH и ФАДH2) окисляются в дыхательной цепи.
И вроде всё просто — давайте перекинем туда НАДH и дело с концом. Пусть он там окисляется, а потом возвращается в цитоплазму. Но тут есть проблема. Во внешней мембране митохондрий есть поры. Через них могу проходить небольшие молекулы, но вот внутренняя мембрана непроницаема. Поэтому НАДH и НАД + не могут проникнуть через неё.
Нужно придумать какой-то механизм, благодаря которому НАДH будет переноситься через внутреннюю мембрану в матрикс митохондрии. Клетка запарилась, но придумала. Её идея ужасно проста, но очень умна (как и всегда). Что если переносить два электрона и протон с НАДH на другую молекулу? Эта молекула должна проходить через одну или две мембраны. Там отдавать электроны и протоны, а потом возвращаться обратно в цитоплазму. И так по кругу.
В общем-то вы уже поняли суть челночных механизмов, а теперь познакомимся с ними поближе. Всего таких механизмов два: глицеролфосфатный и малат-аспартатный.
Глицеролфосфатный челночный механизм
Работает в скелетных мышцах. Давайте посмотрим на общую картинку этого челнока.
Первый этап катализирует цитоплазматическая глицерол-3-фосфатдегидрогеназа. Фермент присоединяет два электрона и два протона к дигидроксиацентофосфату. Образуется глицерол-3-фосфат и НАД + .
Два электрона и протон отдаёт НАДH, а второй протон берётся из цитоплазмы. Но их там не мало. Не запутайтесь в названиях: здесь образуется глицерол-3-фосфат, а не глицеральдегид-3-фосфат из гликолиза.
НАД + остаётся в цитоплазме, где может снова использоваться в шестой реакции гликолиза.
На втором этапе глицерол-3-фосфат проходит через внешнюю мембрану и попадает в межмембранное пространство митохондрии. Там идёт третий этап челночного механизма. Катализирует его митохондриальная глицерол-3-фосфатдегидрогеназа. Фермент находится во внутренней мембране митохондрий. Эта реакция — зеркальное отражение прошлой. Из глицерол-3-фосфата образуется дигидроксиацетонфосфат. Два электрона и два протона принимает кофермент ФАД. Мы получаем дигидроксиацетонфосфат и восстановленный ФАДH2.
Четвёртый этап. Дигидроксиацетонфосфат покидает межмембранное пространство — снова оказывается в цитоплазме. Он опять может повторить цикл.
Вот так мы окислили НАДH до НАД + и перенесли его два электрона и протон в митохондрию. А ещё мы захватили один протон из цитоплазмы, но от неё не убудет. Дальше митохондриальная глицерофосфатдегидрогеназа отдаст два электрона на убихинон, который отправится на третий комплекс дыхательной цепи.
Малат-аспартатный челночный механизм
Работает в большинстве органов и тканей, для нас самые важные — печень и сердце. В нём больше реакций, чем в прошлом челноке. Так что он посложнее.
Первый этап. Электроны и протоны принимает оксалоацетат, который находится в цитоплазме. Катализирует реакцию цитоплазматическая малатдегидрогеназа. Клетка получает окисленный НАД + и малат. НАД + снова идёт в гликолиз. У малата не такая простая судьба.
Малат проходит через внешнюю мембрану митохондрии, но не может пройти через внутреннюю. Для него есть специальный белок переносчик: малат-α-кетоглутарат транслоказа. При её работе малат перемещается в матрикс митохондрий, а альфа-кетоглутарат в межмембранное пространство. Напоминаю, что такой транспорт называется антипортом. Из межмембранного пространства альфа-кетоглутарат может попасть в цитоплазму. Скажу даже больше, он туда попадает…
Второй этап. Малат превращается в оксалоацетат с помощью митохондриальной малатдегидрогеназы. Кофермент этой реакции НАД + , а не ФАД. Образуется НАДH, оксалоацетат и протон. Нужно запомнить, что в малат-аспартатном челноке используется кофермент НАД + .
Теперь нужно вернуть оксалоацетат в цитоплазму, но внутренняя мембрана митохондрии для него непроницаема. Это связано с тем, что он участвует в цикле трикарбоновых кислот. А он как раз идёт в матриксе митохондрий. Поэтому клетка не хочет терять его просто так.
Читайте также: Более двух систем нитей необходимо для выработки тканей переплетения
Третий этап. Для обхода этого защитного механизма есть дополнительная реакция. Эту реакцию катализирует митохондриальная аспартатаминотрансфераза. Фермент переносит аминогруппу с глутамата на оксалоацетат, для этого ему нужен кофермент — пиридоксальфосфат. Образуется аспартат и альфа-кетоглутарат.
Аспартат переносится через внутреннюю мембрану с помощью белка переносчика — глутамат-аспартат транслоказы. Она переносит аспартат в межмембранное пространство митохондрий, а глутамат в матрикс. Там глутамат используется для трансаминирования — мы видели это выше.
Четвёртый этап. Снова превращаем аспартат в оксалоацетат. Здесь нам понадобится альфа-кетоглутарат, который мы переносили в цитоплазму на первом этапе. Катализирует реакцию цитоплазматическая аспартатаминотрансфераза. Снова образуется оксалоацетат и глутамат. Оксалоацетат используется для малат-аспартатного челночного механизма, а глутамат переносится в митохондрию.
Цикл завершён. Мы перенесли электроны и протон с НАДH в матрикс митохондрии.
В челноках было много одинаковых по функциям ферментов. Но при этом у каждого была приписочка: цитоплазматический или митохондриальный. Дело в том, что это изоферменты — они выполняют одинаковую функцию, но при этом отличаются по аминокислотным последовательностям. А это значит, что они кодируются разными генами.
С аспартатаминотрансферазой (АСТ) вы встретитесь ещё миллиард раз. Она используется при диагностике инфаркта миокарда и заболеваний печени. А всё почему? Потому что там наибольшая активность малат-аспартатного челнока. Так что запоминайте этот фермент и не думайте, что биохимия бесполезна. Опять говорю как препод….
Что происходит с электронами дальше?
Смысл окисления НАДH до НАД + не только в бесперебойной работе гликолиза. Электроны и протоны, которые были перенесены в матрикс митохондрий используются для синтеза АТФ. Как? С помощью дыхательной цепи. Я не буду расписывать её ещё раз, если хотите подробнее, то о ней можно почитать тут. Главное сейчас — это понять суть.
Электроны переносятся от одного комплекса дыхательной цепи к другому, пока они не попадут на кислород. В ходе такого переноса выделяется энергия. Эта энергия используется для транспорта протонов водорода из матрикса митохондрий в межмембранное пространство. Так создаётся градиент концентрации. Внутри митохондрии протонов мало, а снаружи много. При этом первый комплекс выкачивает четыре протона, третий тоже четыре, а четвёртый только два протона. Градиент концентрации используется на синтез АТФ с помощью АТФ-синтазы. На синтез одной молекулы АТФ затрачивается
В глицеролфосфатном челноке мы получили ФАДH2. Электроны с него попадают сразу на убихинон, то есть пропускают первый комплекс дыхательной цепи. Значит в ходе путешествия электронов по дыхательной цепи мы получим всего шесть протонов водорода. Или 1,5 молекулы АТФ — на синтез одной молекулы нужно четыре протона. Это из одного ФАДH2. Но в ходе гликолиза образовались две молекулы НАДH, а значит мы получим две молекулы ФАДH2. Умножим 1,5 на 2 и получим три молекулы АТФ — из каждого ФАДH2 по 1,5 АТФ. Прибавим две, которые мы получили в ходе гликолиза. И выйдет пять молекул АТФ. Это общий выход гликолиза при использовании глицеролфосфатного челнока.
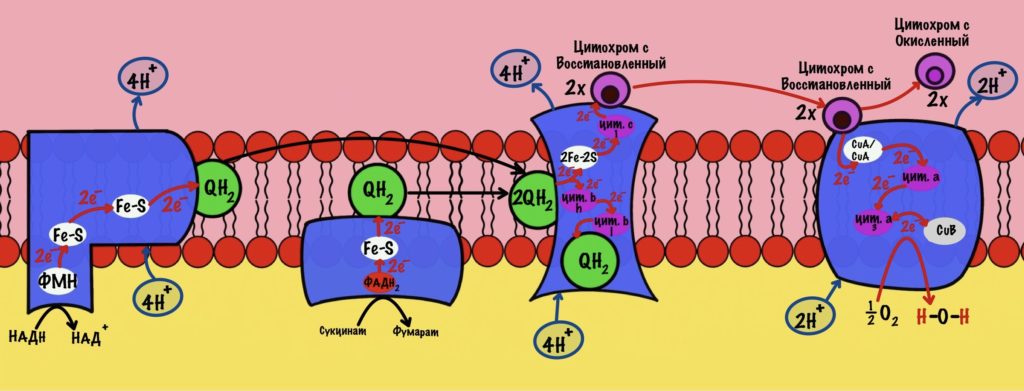
В ходе малат-аспартатного челнока был получен НАДH. Он отдаёт электроны на первый комплекс дыхательной цепи. Значит общее количество протонов — десять. Или 2,5 молекулы АТФ. В ходе гликолиза мы получили два НАДH, а значит и здесь их будет два. Общее количество АТФ — пять. Прибавим две, которые мы получили в ходе гликолиза. Выходит семь молекул АТФ. Это общий выход гликолиза при использовании малат-аспартатного челнока.
Хочешь задать вопрос, похвалить или наговорить гадостей? Тогда залетай в телегу. Там ты сможешь предложить новый формат или разбор темы. А если серьёзно, то эти статьи пишутся для вас, поэтому мне важна обратная связь.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом