
Ключевыми ферментами этого челнока являются изоферменты малатдегидрогеназы – цитоплазматический и митохондриальный. Он является распространенным по всем тканям.
Этот механизм более сложен: постоянно идущие в цитоплазме при участии фермента аспартатаминотрансферазы (АСТ) реакции трансаминированияаспарагиновой кислоты с α-кетоглутаратом поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназыи за счет «гликолитического» НАДН восстанавливается до яблочной кислоты(малата).
Последняя антипортом с α-кетоглутаратом проникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он при помощи аспартатаминотрансферазы трансаминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль.
Таким образом, атомы водорода от цитозольного НАДН перемещаются в состав митохондриального НАДН.

Схема работы малат-аспартатной челночной системы
Организм приспособлен к обезвреживанию этанола
В организм поступают и в результате метаболизма образуются в клетках головного мозга и печени, в других тканях, при жизнедеятельности микрофлоры кишечника различные спирты (алифатической, ароматической, стероидной природы, ретиноиды, фарнезол и др.) и альдегиды (ароматические, алифатические, продукты пероксидации липидов и др.), которые могут являться промежуточными метаболитами или конечными продуктами. Взаимопревращение спиртов и альдегидов осуществляют алкогольдегидрогеназы. Их существует 6 классов, в каждом классе имеются многочисленные изоферменты, обнаруженные во многих тканях.
С медицинской и социальной точки зрения определенный интерес вызывает метаболизм этилового спирта в организме человека.
Обезвреживание этанола
Метаболизм поступающего этанола в организме происходит преимущественно в печени двумя путями. Первый путь заключается в окислении спирта по алкогольдегидрогеназному пути до уксусной кислоты, которая в виде ацетил-SКоА поступает в ЦТК. Через этот путь проходит от 70% до 90% всего этанола.

Реакции окисления этанола и ацетальдегида
Оставшаяся часть окисляется в эндоплазматическом ретикулуме (микросомальное окисление) алкогольоксидазой. При регулярном поступлении этанола доля микросомального окисления возрастает, количество молекул алкогольоксидазы увеличивается.
Побочные эффекты обезвреживания этанола
Поскольку при утилизации этанола образуется большое количество НАДН, то в цитозоле гепатоцитов активируется 11-я реакция гликолиза – превращение пирувата в лактат и восстановление диоксиацетонфосфата в глицерол-3-фосфат. Одновременно накопление «алкогольного» ацетил-SКоА ингибирует пируватдегидрогеназу, что еще больше усиливает накопление лактата.
Все это приводит к гипогликемии так как пировиноградная кислота и диоксиацетонфосфат являются субстратами глюконеогенеза. Свободный выход молочной кислоты в кровь обуславливает лактатацидемию (лактоацидоз).
Одновременно из-за относительной недостаточности оксалоацетата избыток «алкогольного» ацетил-SКоА не успевает окислиться в цикле трикарбоновых кислот и перенаправляется на синтез кетоновых тел, что обеспечивает возникновение кетоацидоза.
Спиртовое брожение
Образование этилового спирта из глюкозы происходит в анаэробных условиях в дрожжах и некоторых видах плесневых грибков. Суммарное уравнение реакции:
До стадии образования пирувата реакции спиртового брожения совпадают с реакциями гликолиза. Отличия заключаются только в дальнейшем превращении пировиноградной кислоты. Цель этих превращений у указанных организмов та же, что и в гликолизе (молочнокислом брожении) – удалить пируват из клетки и окислить НАДН, который образовался в 6-й реакции.
Читайте также: Эмбриональное происхождение хрящевой ткани
Малат-аспартатный челночный механизм
Ключевыми ферментами этого челнока являются изоферменты малатдегидрогеназы – цитоплазматический и митохондриальный. Он является распространенным по всем тканям.
Этот механизм более сложен: постоянно идущие в цитоплазме при участии фермента аспартатаминотрансферазы (АСТ) реакции трансаминированияаспарагиновой кислоты с α-кетоглутаратом поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназыи за счет «гликолитического» НАДН восстанавливается дояблочной кислоты(малата).
Последняя антипортом с α-кетоглутаратом проникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он при помощи аспартатаминотрансферазы трансаминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль.
Таким образом, атомы водорода от цитозольного НАДН перемещаются в составмитохондриального НАДН.
Гликоген — (C6H10O5)n, полисахарид, образованный остатками глюкозы, связанными α-1→4связями (α-1→6 в местах разветвления); основной запасной углевод животных. Гликоген является основной формой хранения глюкозы в животных клетках. Откладывается в виде гранул вцитоплазме во многих типах клеток (главным образом печени и мышц).
Гликоген иногда называется животным крахмалом, так как его структура похожа наамилопектин — компонент растительного крахмала. В отличие от крахмала, гликоген имеет более разветвленную и компактную структуру, не дает синей окраски при окраске йодом.
Гликоген образует энергетический резерв, который может быть быстро мобилизован при необходимости восполнить внезапный недостаток глюкозы. Гликогеновый запас, однако, не столь ёмок в калориях на грамм, как запас триглицеридов (жиров). Только гликоген, запасённый в клетках печени (гепатоциты) может быть переработан в глюкозу для питания всего организма. Содержание гликогена в печени при увеличении его синтеза может составить 5-6% от массы печени [1] . Общая масса гликогена в печени может достигать 100—120 граммов у взрослых. В мышцах гликоген перерабатывается в глюкозу исключительно для локального потребления и накапливается в гораздо меньших концентрациях (не более 1 % от общей массы мышц), в то же время его общий мышечный запас может превышать запас, накопленный в гепатоцитах. Небольшое количество гликогена обнаружено в почках, и ещё меньшее — в определённых видах клеток мозга (глиальных) и белых кровяных клетках.
В качестве запасного углевода гликоген присутствует также в клетках грибов.
группа ферментов класса гидролаз, катализирующихгидролитическое расщепление эфирных связей в гликозидах; участвуют в процессах превращенияуглеводов.
Гликозил-гидролазы (или гликозидазы) катализируют гидролиз гликозидных связей в молекулах углеводов, приводя к появлению двух более мелких молекул углеводов. Эти ферменты встречаются в клетках почти всех живыхорганизмов. Гликозидазы выполняют множество разнообразных функций: деградация биомассы (например,целлюлозы и гемицеллюлозы), участие в антибактериальной защите организма (например, лизоцим), развитиепатогенеза (например, вирусные нейраминидазы), клеточный биосинтез (например, маннозидазы, вовлечённые в созревание N-гликозилированных гликопротеинов). Гликозидазы вместе с гликозилтрансферазами образуют основу биологического аппарата синтеза и разрушения гликозидных связей.
Читайте также: Характеристика тканей человека примеры
Эндергонические и экзергонические реакции
Направление химической реакции определяется значением ΔG. Если эта величина отрицательна, то реакция протекает самопроизвольно и сопровождается уменьшением свободной энергии. Такие реакции называют экзергоническими. Если при этом абсолютное значение ΔG велико, то реакция идёт практически до конца, и её можно рассматривать как необратимую.
Если ΔG положительно, то реакция будет протекать только при поступлении свободной энергии извне; такие реакции называют эндергоническими.
Если абсолютное значение ΔG велико, то система устойчива, и реакция в таком случае практически не осуществляется. При ΔG, равном нулю, система находится в равновесии

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).


Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ — конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.
Малат-аспартатный челнок
Трансфер малат-аспартат является механизмом для транспортировки молекул в эукариот через внутреннюю мембрану митохондрий в целях передачи NADH + H + из гликолиза , полученного в цитозоле к митохондриальной матрице за счет регенерации NAD + в цитозоле.
Операция
Поскольку внутренняя митохондриальная мембрана непроницаема для коферментов, таких как НАДН, она пересекает эту мембрану косвенно, через обмен четырьмя метаболитами, который стал возможным благодаря системе из четырех ферментов — два в цитозоле и две изоформы предыдущих в матриксе митохондрий. — и два антипорта ( пермеаса ) внутренней митохондриальной мембраны:
- НАДН передает два электрона к молекуле из оксалоацетатацитозольного под действием малатдегидрогеназы цитозола (CMDh);
- полученный цитозольный малат может пересекать внешнюю митохондриальную мембрану с использованием порина, но должен заимствовать малат- α-кетоглутаратантипорт, чтобы пересечь внутреннюю митохондриальную мембрану ;
- малата , который достиг митохондриального матрикса является окисленным снова оксалоацетат митохондриальной малатдегидрогеназы (mMDH) , чтобы преобразовать молекулу NAD + в NADH;
- оксалоацетат превращаются в аспартаты под действием митохондриальной аспартатаминотрансферазы (тучная) путем передачи в аминныхгруппы из глутамата преобразованы в альфа-кетоглутарат ;
- аспартат заимствует глутамат-аспартатный антипорт для пересечения внутренней митохондриальной мембраны, затем пересекает внешнюю митохондриальную мембрану через порин, чтобы восстановить цитозоль;
- Цитозольный аспартат превращается в оксалоацетат цитозольной аспартатаминотрансферазой (cAST), которая также производит глутамат из α-кетоглутарата .
Преимущество этого механизма заключается в сохранении потенциала переноса электронов, производимых цитозольным окислением, поскольку эти электроны передаются косвенно от цитозольного НАДН к митохондриальному НАДН.
Читайте также: Чем почистить ткань дверной карты
Следовательно, это не активный транспорт , поэтому он эффективен только тогда, когда концентрация цитозольного НАДН значительно выше, чем концентрация митохондриального матрикса. У млекопитающих этот челнок в основном воздействует на сердце , печень и почки .
Ионов кальция Са 2+ , вторичные мессенджеры из мышечных сокращений , действуют как активаторы этого челнока.
Метаболическая роль
С другой стороны, есть еще один митохондриальный челнок , глицерин-3-фосфатный челнок , который потребляет часть потенциала переноса электронов цитозольного НАДН — поэтому он немного менее энергетически эффективен — но быстрее и позволяет высвобождать l энергия раньше потребляла такие органы, как мышцы и мозг .
Малат-аспартатный челнок позволяет производить около 2,5 молекул АТФ на цитозольный НАДН, в то время как глицерин-3-фосфатный челнок позволяет производить около 1,5 молекул АТФ на цитозольный НАДН.
ЧЕЛНОЧНЫЕ МЕХАНИЗМЫ
Цитозольный НАДH, образующийся в 6 реакции гликолиза, катализируемой глицеральдегидфосфатдегидрогеназой, не может передавать водород в дыхательную цепь, потому что митохондриальная мембрана для него непроницаема. Перенос водорода через мембрану происходит с помощью специальных систем, называемых «челночными». Водород транспортируется через мембрану при участии пар субстратов, связанных соответствующими дегидрогеназами, т.е. с обеих сторон митохондриальной мембраны находится специфическая дегидрогеназа.
Глицеролфосфатная челночная системаработает в клетках белых мышц, печени, мозге.
Водород от НАДH в цитозоле передается на диоксиацетонфосфат глицерол-3-фосфатдегидрогеназой (НАД-зависимый фермент) (рис. 38). Образовавшийся глицерол-3-фосфат окисляется ферментом внутренней мембраны митохондрий — глицерол-3-фосфатдегидрогеназой (ФАД-зависимый фермент). Затем протоны и электроны с ФАДH2 переходят на убихинон и далее по дыхательной цепи.
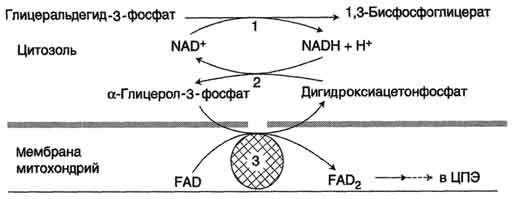
Рис. 38. Глицеролфосфатная челоночная система:
2 — глицерол-3-фосфатдегидрогеназа (цитозольный фермент);
3 — глицерол-3-фосфатдегидрогеназа (митохондриальныи фермент)
Малат-аспартатная челночная система, в которой участвуют малат, цитозольная и митохондриальная малат-дегидрогеназы, является более универсальной, и работает в сердечной мышце, печени, почках.
В цитоплазме НАДH восстанавливает оксалоацетат в малат (реакция 1), который при участии переносчика проходит в митохондрии, где окисляется в оксалоацетат НАД-зависимой малатдегидрогеназой (реакция 2). Восстановленный НАДН отдает водород в митохондриальную ЦПЭ (рис. 39).
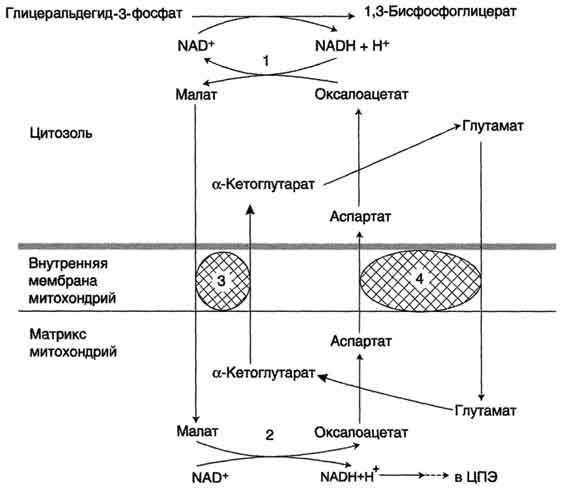
Рис. 39. Малат-аспартатная челоночная система:
1, 2 — окислительно-восстановительные реакции, обеспечивающие транспорт водорода из цитозоля в митохондрии на ЦПЭ;
3, 4 — транслоказы, обеспечивающие транспорт a-кетоглутарата, аспартата и глутамата и через мембрану митохондрий
Образованный из малата оксалоацетат выйти из митохондрий в цитозоль не может: мембрана митохондрий для него непроницаема. Поэтому он превращается в аспартат, который транспортируется в цитозоль, где снова превращается в оксалоацетат.
Обе челночные системы отличаются по количеству синтезированной АТФ. В первой системе образуется 2АТФ (водород вводится в ЦПЭ на уровне убихинона). Вторая система энергетически более эффективна – образуется 3АТФ (водород поступает в ЦПЭ с митохондриальным НАД + ).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом