
В какую ткань инсулин увеличивает транспорт глюкозы
а) Влияние инсулина на обмен углеводов. Сразу после потребления пищи, богатой углеводами, глюкоза, всасываясь в кровь, стимулирует секрецию инсулина. Инсулин, в свою очередь, повышает поступление, хранение и использование глюкозы почти всеми тканями организма, особенно мышцами, жировой тканью и печенью.
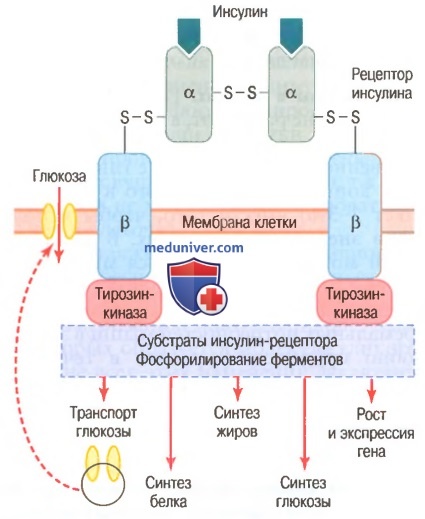
Схема инсулин-рецептора. Инсулин связывается с альфа-субъединицей рецептора, что вызывает фосфорилирование бета-субъединицы рецептора.
Это, в свою очередь, индуцирует активность тирозинкиназы. Рецептор тирозинкиназной активности запускает каскад реакций фосфорилирования в клетке, что увеличивает или снижает активность ферментов, включая субстраты инсулин-рецептора, которыми опосредованы влияния глюкозы на метаболизм глюкозы, жиров и белков.
Например, транспортеры глюкозы продвигаются к мембране клетки, облегчая поступление глюкозы в клетку
б) Инсулин повышает поступление и метаболизм глюкозы в мышцах. Неоднократно на протяжении суток мышечная ткань обеспечивает свои энергетические потребности не столько глюкозой, сколько жирными кислотами. Это объясняется тем, что мембрана мышечной клетки в покое обладает низкой проницаемостью для глюкозы, кроме случаев, опосредованных стимулирующими влияниями инсулина; в промежутках между приемами пищи количество секретируемого инсулина слишком мало, чтобы обеспечить поступление значительного количества глюкозы в клетки.
Однако существуют два состояния, когда мышцы потребляют большое количество глюкозы. Одно из них — тяжелые физические нагрузки. Потребление глюкозы в такой ситуации не требует большого количества инсулина, поскольку мышечные волокна становятся более проницаемы для глюкозы даже при отсутствии инсулина за счет процессов сокращения.
Вторым случаем высокого потребления глюкозы мышцами становится период через несколько часов после приема пищи. В это время концентрация глюкозы в крови высока и поджелудочная железа секретирует большое количество инсулина. Высокая концентрация инсулина обеспечивает быстрый транспорт глюкозы в мышечные клетки. Это вызывает преимущественное по сравнению с жирными кислотами использование глюкозы мышечными клетками, что мы обсудим далее.
в) Хранение глюкозы в мышцах. Если мышцы после приема пищи не работают, а глюкоза в больших количествах еще поступает в клетки, она преимущественно запасается в виде гликогена (предельная концентрация которого в мышцах составляет 2-3%) вместо того, чтобы использоваться в энергетических целях. В дальнейшем гликоген может использоваться мышцами для получения энергии.
Депонирование гликогена особенно полезно во время коротких периодов экстремальных потребностей мышц в энергии и способно продемонстрировать в течение нескольких минут высокий уровень энергообеспечения даже при отсутствии кислорода за счет гликолитического расщепления гликогена до молочной кислоты.
г) Количественное влияние инсулина на облегчение транспорта глюкозы через мембраны мышечных клеток. Это влияние показано на рисунке ниже в виде результатов экспериментов.
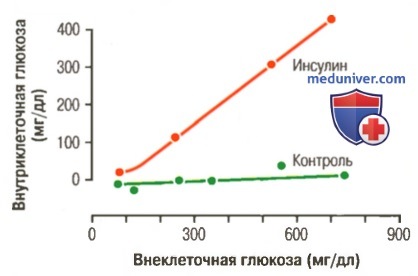
Влияние инсулина, облегчающее повышение концентрации глюкозы внутри мышечных клеток.
Заметьте, что при отсутствии инсулина (контроль) внутриклеточная концентрация глюкозы остается близкой к нулю, несмотря на ее высокую внеклеточную концентрацию
Нижняя кривая («контроль») показывает концентрацию свободной глюкозы, измеренную внутри клетки. Концентрация свободной глюкозы остается почти равной нулю, несмотря на то, что концентрация глюкозы во внеклеточной жидкости достигает 750 мг/дл. Кривая, помеченная «инсулин», показывает, что концентрация глюкозы во внутриклеточной жидкости достигает 400 мг/дл при добавлении инсулина. Таким образом, инсулин может повышать скорость транспорта глюкозы в покоящиеся мышечные клетки почти в 15 раз.
Видео физиология гормонов поджелудочной железы и регуляции уровня глюкозы — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
В какую ткань инсулин увеличивает транспорт глюкозы
• Для того чтобы преодолеть гематоэнцефалический барьер, глюкоза транспортируется в астроциты через эндотелиальные клетки небольших кровеносных сосудов
Читайте также: Как сделать шляпку цветка из ткани
• Белки переносят глюкозу за счет унипорта, транспортируя ее в направлении градиента концентрации
• Белки, переносящие глюкозу, претерпевают конформационные изменения, которые приводят к реориентации их субстратных сайтов в мембране клетки
В клетках эукариот основным источником энергии является глюкоза, и многие клетки нуждаются в постоянном ее поступлении, поскольку у них глюкоза служит основным источником энергии для синтеза АТФ. Глюкоза представляет собой полярную молекулу, которая способна к гидратации, а клеточные мембраны не пропускают такие небольшие полярные метаболиты, как сахара. Поэтому для транспорта глюкозы в клетку требуется участие специфических мембранных белков. Транспорт глюкозы через плазматическую мембрану происходит с участием продуктов двух семейств генов.
Переносчики глюкозы (GLUTs) осуществляют независимый перенос (унипорт), при котором через мембрану происходит облегченный транспорт глюкозы. В противоположность GLUT-белкам, котранспортеры Na+/глюкоза расходуют энергию трансмембранного градиента Na+ на транспорт глюкозы. В настоящем разделе мы рассмотрим GLUT-белки.
![]()
Глюкоза селективно транспортируется через гематоэнцефалический барьер посредством изоформы 1 (GLUT-1) белка-переносчика.
Перенос глюкозы из крови в головной мозг и в другие отделы ЦНС представляет собой многоступенчатый процесс,
в котором участвуют различные типы клеток.
Семейство GLUT-белков является частью более обширного суперсемейства белков, облегчающих основные транспортные процессы (MFS) и широко представленных в клетках всех организмов. GLUT-белки представляют собой интегральные белки мембран клеток эукариот. Изоформы переносчиков GLUT различаются по своим кинетическим характеристикам, специфичностью по отношению к переносимым сахарам, тканевой локализацией и механизмами регуляции. Некоторые GLUT-белки, наряду с глюкозой, транспортируют и другие метаболиты, например галактозу, воду и анальгетики из группы гликопептидов.
GLUT-белки осуществляют унипорт, при котором вещества проходят через мембрану в направлении градиента концентрации. Таким образом, в зависимости от концентрации, GLUT-белки переносят метаболиты в клетку или в противоположном направлении. Поступление в клетку глюкозы, происходящее при участии GLUT-белков, часто определяет жизнеспособность клеток, которые характеризуются высоким уровнем потребления энергии.
Питательные метаболиты, такие как сахара, через кровеносные сосуды транспортируются в органы. Эндотелиальные клетки, выстилающие стенки небольших сосудов, контролируют процессы обмена питательных веществ. В этих эндотелиальных клетках, особенно расположенных в области гематоэнцефалического барьера, содержится много GLUT-белков. Надлежащее функционирование головного мозга сильно зависит от глюкозы, и его клетки особенно чувствительны к снижению ее содержания.
Транспорт глюкозы в нервные клетки происходит через капилляры мозга в несколько этапов и с участием изоформы GLUT-1. Эта изоформа экспрессируется в мембране клеток эндотелия, находящихся на границе между кровью и межклеточным пространством, а также в плазматической мембране астроцитов, функция которых важна в гематоэнцефалическом барьере. Расположенные в этих местах белки GLUT-1 транспортируют глюкозу из крови в эндотелиальные клетки, и оттуда в астроциты. В них глюкоза превращается в другие источники энергии, которые транспортируются в нейроны.
В различных тканях содержатся разные изоформы GLUT. Например, в клетках мышечной и жировой ткани транспорт глюкозы осуществляется с участием GLUT-4. В процессе приема пищи или после еды в клетки этих тканей под действием инсулина увеличивается поступление глюкозы. При этом происходит регулируемый транспорт изоформы GLUT-4 к клеточной поверхности. Эта изоформа также называется инсулин-зависимый переносчик. Белок GLUT-4 локализован во внутриклеточных везикулах, которые сливаются с плазматической мембраной. Этим обеспечивается доставка переносчика GLUT-4 к плазматической мембране и увеличивается емкость транспортного процесса.
![]()
Предполагаемое строение GLUT-переносчика, состоящего из 12 трансмембранных сегментов с внутриклеточными С- и N-концевыми участками.
Внутриклеточные петли содержат сайты фосфорилирования и связывания субстратов.
Показано, что градиент глюкозы по обеим сторонам мембраны может возникать в любом направлении, в зависимости от типа клеток и их метаболизма.
Градиент определяет направление транспорта.
Читайте также: Как называется принтер для ткани
Связывание инсулина со своим поверхностным рецептором запускает каскад внутриклеточных процессов, приводящих к быстрому слиянию этих везикул с мембраной. Это, в свою очередь, приводит к быстрому увеличению транспорта глюкозы в клетку с участием GLUT-4. При диабете типа II не происходит транспорт глюкозы из крови в мышцы и жировую ткань, очевидно, из-за нарушения позиционирования GLUT-4 на плазматической мембране. Более того, GLUT-2 способен экспортировать глюкозу из клеток тех органов, в которых она образуется, например из клеток печени.
По строению переносчик GLUT напоминает другие члены суперсемейства MFS. Предполагается, что он состоит из 12 трансмембранных а-спиралей, имеющих внутриклеточные N- и С-концевые участки и петли. Последние содержат сайт связывания субстрата и сайты фосфорилирования.
Модель структуры GLUT-1 предложена на основании результатов по сайт-направленному мутагенезу и по измерению транспорта глюкозы мутантным белком. Для построения модели также использовались данные по кристаллической структуре бактериальной лактопермеазы, относящейся к подсемейству переносчиков олигосахари-дов/Н+, входящих в MFS.
Согласно этой модели, при соответствующей ориентации трансмембранных спиралей создается полость поры, через которую транспортируется глюкоза, а также обеспечивается образование водородных связей между GLUT-белком и глюкозой. Результаты кинетического анализа транспорта глюкозы в эритроцитах позволяют предполагать существование специального механизма, который обеспечивает белку GLUT возможность принимать одну из двух основных конформаций. Этот механизм аналогичен постулируемому для бактериальной лактопермеазы. Когда белок находится в одной конформации, сайт связывания глюкозы обращен в сторону внеклеточного пространства. В другой конформации он обращен в сторону цитозоля. При связывании глюкозы с любой стороны происходят конформационные изменения, которые приводят к реориентации сайтов связывания глюкозы к противоположной стороне мембраны и к ее высвобождению.
Таким образом, хотя переносчики GLUT являются унипортерами, а бактериальная лактопермеаза представляет собой симпортер, предполагается, что они функционируют по одному механизму.
Мутации в гене GLUT-1 вызывают значительные дефекты развития. У детей потребность головного мозга в глюкозе в 3-4 раза выше, чем у взрослых, и в мозг поступает до 80% всей глюкозы. Мутации в гене GLUT-1 человека служат причиной проявления редкого синдрома недостаточности GLUT-1, который характеризуется развитием припадков и замедлением развития. Считается, что это связано с нарушением транспорта глюкозы в головной мозг. Эмбрионы мышей, дефектных по гену GLUT-1, отстают в росте, и у них обнаруживаются различные пороки развития. Аналогичные дефекты возникают у мышиных эмбрионов, которые развиваются в организме диабетических самок. При этом избыток глюкозы в крови подавляет экспрессию GLUT-1 в органах эмбриона.
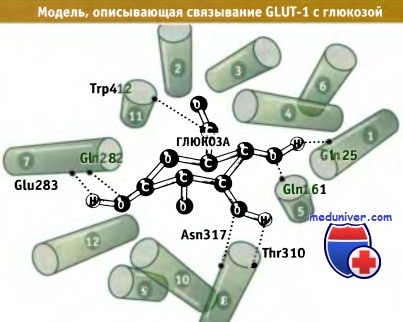
Предполагаемая ориентация трансмембранных а-спиралей переносчика глюкозы GLUT-1.
Показаны остатки, участвующие в связывании глюкозы (размер молекулы глюкозы представлен без учета масштаба).
Вид на спирали со стороны внутренней части плазматической мембраны.
Эта модель построена с использованием гомологичной структуры лактопермеазы E.coli в качестве матрицы.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
В какую ткань инсулин увеличивает транспорт глюкозы
Действие инсулина на клетки-мишени начинается после его связывания со специфическими димерными мембранными рецепторами (рис. 6.22), при этом внутриклеточный домен рецептора обладает тирозинкиназной активностью. Инсулин-рецепторный комплекс не только передает сигнал внутрь клетки, но и частично путем эндоцитоза поступит внутрь клетки к лизо-сомам. Под влиянием лизосомальной протеазы инсулин отщепляется от рецептора, при этом последний либо разрушается, либо возвращается к мембране и вновь встраивается в нее. Многократное перемещение рецептора от мембраны к лизосомам и обратно к мембране носит название рециклизация рецептора. Процесс рециклизации важен для регуляции колич-ства инсулиновых рецепторов, в частности обеспечения обратной зависимости между концентрацией инсулина и количеством мембранных рецепторов к нему.
Читайте также: Мукозоассоциированная лимфоидная ткань это что
Образование инсулин-рецепторного комплекса активирует тирозинкиназу, запускающую процессы фосфорилирования внутриклеточных белков. Происходящее при этом аутофосфорилирование рецептора ведет к усилению первичного сигнала. Инсулин-рецепторный комплекс вызывает активирование фосфолипазы С, образование вторичных посредников инозитолтрифосфата и диацилглицерола, активацию протеинкиназы С, ингибирование цАМФ. Участие нескольких систем вторичных посредников объясняет многообразие и различия эффектов инсулина в разных тканях.
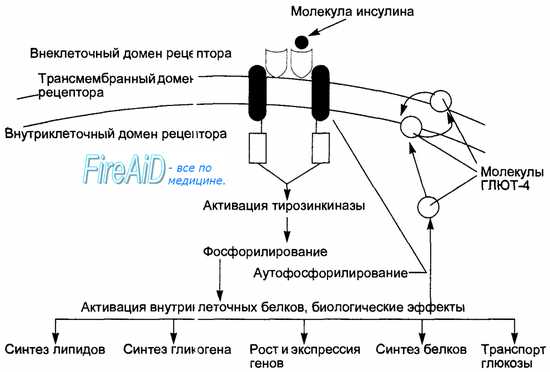
Рис. 6.22. Схема механизма действия инсулина на клетку-мишень.
Инсулин оказывает влияние на все виды обмена веществ, способствует анаболическим процессам, увеличивая синтез гликогена, жиров и белков, тормозя эффекты многочисленных контринсулярных гормонов (глюкагона, катехоламинов, глюкокортикоидов и соматотропина). Все эффекты инсулина по скорости их реализации подразделяют на 4 группы: очень быстрые (через несколько секунд) — гиперполяризация мембран клеток (за исключением гепатоцитов), повышение проницаемости для глюкозы, активация Na-K-АТФазы, входа К+ и откачивания Na , подавление Са-насоса и задержка Са2+; быстрые эффекты (в течение нес кольких минут) — активация и торможение различных ферментов, подавляющих катаболизм и усиливающих анаболические процессы; медленные процессы (в течение нескольких часов) — повышенное поглощение амиминокислот, изменение синтеза РНК и белков-ферментов; очень медленные эффекты (от часов до суток) — активация митогенеза и размножения клеток.
Важнейшим эффектом инсулина в организме является увеличение в 20—50 раз транспорта глюкозы через мембраны мышечных и жировых клеток путем облегченной диффузии по градиенту концентрации с помощью чувствительных к гормон) мембранных белковых переносчиков, называемых ГЛЮТ. В мембранах разных видов клеток выявлены 6 типов ГЛЮТ (рис. 6.23), но только один из них — ГЛЮТ-4 — является инсулинозависимым и находится в мембранах клеток скелетных мышц, миокарда, жировой ткани.
Инсулин влияет на угле водный обмен, что проявляется:
1) активацией утилизации глюкозы клетками,
2) усилением процессов фосфорилирования;
3) подавлением распад; и стимуляцией синтеза гликогена;
4) угнетением глюконеогенеза;
5) активацией процессов гликолиза;
6) гипогликемией.
Действие инсулина на белковый обмен состоит в: 1) повышении проницаемости мембран для аминоокислот; 2) усилении синтеза иРНК; 3) активации в печени синтеза aминокислот; 4) повышении синтеза и подавлении распада белка.
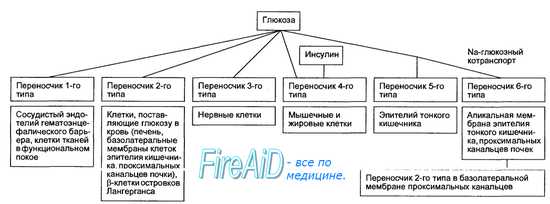
Рис. 6.23. Схема транспорта глюкозы через клеточные мембраны. Переносчики имеют общее название ГЛЮТ-1, 2, 3, 4, 5, 6. Только ГЛЮТ-4 является инсулинозависимым.
Основные эффекты инсулина на липидный обмен:
• стимуляция синтеза свободных жирных кислот из глюкозы;
• стимуляция синтеза липопротеиновой липазы в клетках эндотелия сосудов и благодаря этому активация гидролиза связанных с липо-протеинами крови триглицеридов и поступления жирных кислот в клетки жировой ткани;
• стимуляция синтеза триглицеридов;
• подавление распада жира;
• активация окисления кетоновых тел в печени.
Благодаря влиянию на клеточную мембрану инсулин поддерживает высокую внутриклеточную концентрацию ионов калия, что необходимо для обеспечения нормальной в возбудимости клеток.
Широкий спектр метаболических эффектов инсулина в организме свидетельствует о том, что гормон необходим для осуществления функционирования всех тканей, оргаганов и физиологических систем, реализации эмоциональных и поведенческих актов, поддержания гомеостазиса, осуществления механизмов приспособления и защиты организма от неблагоприятных факторов среды.
Недостаток инсулина (относительный дефицит по сравнению с уровнем контринсулярных гормонов, прежде всего глюкагона) приводит к сахарному диабету. Избыток инсулина в крови, например при передозировке, вызывает гипогликемию с резкими нарушениями функций центральной нервной системы, использующей глюкозу как основной источник энергии независимо от инсулина.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом