
Величина возбудимости ткани в фазу быстрой деполяризации потенциала действия равна
В нервных волокнах сигналы передаются с помощью потенциалов действия, которые представляют собой быстрые изменения мембранного потенциала, быстро распространяющиеся вдоль мембраны нервного волокна. Каждый потенциал действия начинается со стремительного сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу. При проведении нервного сигнала потенциал действия движется вдоль нервного волокна вплоть до его окончания.
На рисунке показаны изменения, возникающие на мембране во время потенциала действия, с переносом положительных зарядов внутрь волокна вначале и возвращением положительных зарядов наружу в конце. В нижней части рисунка графически представлены последовательные изменения мембранного потенциала в течение нескольких 1/10000 сек, иллюстрирующие взрывное начало потенциала действия и почти столь же быстрое восстановление.
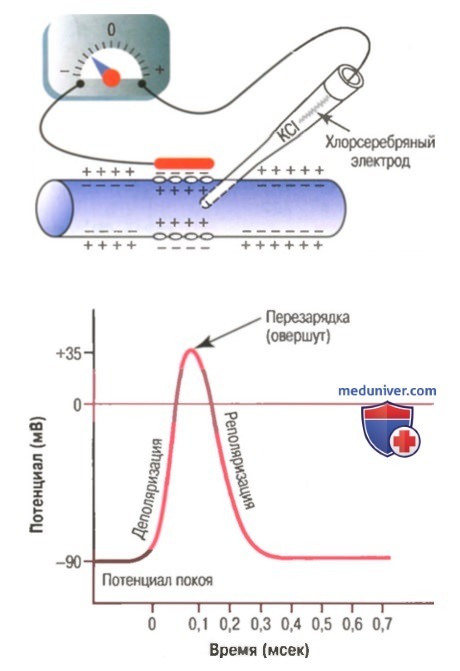
Типичный потенциал действия, зарегистрированный с помощью метода, представленного в верхней части рисунка.
Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала, равного -90 мВ.
Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь аксона. Нормальное поляризованное состояние в -90 мВ немедленно нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении. Этот процесс называют деполяризацией, В крупных нервных волокнах значительный избыток входящих внутрь положительных ионов натрия обычно приводит к тому, что мембранный потенциал «проскакивает» за пределы нулевого уровня, становясь слегка положительным. В некоторых более мелких волокнах, как и в большинстве нейронов центральной нервной системы, потенциал достигает нулевого уровня, не «перескакивая» его.
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Для более полного понимания факторов, являющихся причиной деполяризации и реполяризации, необходимо изучить особенности двух других типов транспортных каналов в мембране нервного волокна: электроуправляемых натриевых и калиевых каналов.
Электроупавляемые натриевые и калиевые каналы. Необходимым участником процессов деполяризации и реполяризации во время развития потенциала действия в мембране нервного волокна является электроуправляемый натриевый канал. Электроуправляемый калиевый канал также играет важную роль в увеличении скорости реполяризации мембраны. Оба типа электроуправляемых каналов существуют дополнительно к Na + /K + -насосу и каналам К + /Na + -утечки.
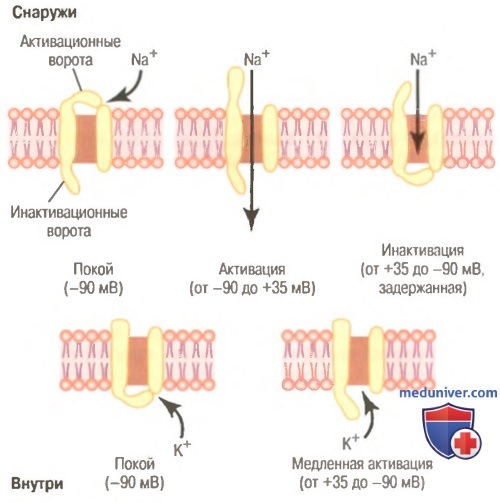
Типичные изменения состояния электроуправляемых натриевых (вверху) и калиевых (внизу) каналов при изменении мембранного потенциала от нормального отрицательного значения до положительных величин.
Показана последовательная активация и инактивация натриевых каналов и задержанная активация калиевых каналов.
Электроуправляемый натриевый канал. В верхней части рисунка показан электроуправляемый натриевый канал в трех различных состояниях. Этот канал имеет двое ворот: одни вблизи наружной части канала, которые называют активационными воротами, другие — у внутренней части канала, которые называют инактивационными воротами. В верхней левой части рисунка изображено состояние этих ворот в покое, когда мембранный потенциал покоя равен -90 мВ. В этих условиях активационные ворота закрыты и препятствуют поступлению ионов натрия внутрь волокна.
Активация натриевого канала. Когда мембранный потенциал покоя смещается в направлении менее отрицательных значений, поднимаясь от -90 мВ в сторону нуля, на определенном уровне (обычно между -70 и -50 мВ) происходит внезапное конформационное изменение актива-ционных ворот, в результате они переходят в полностью открытое состояние. Это состояние называют активированным состоянием канала, при котором ионы натрия могут свободно входить через него внутрь волокна; при этом натриевая проницаемость мембраны возрастает в диапазоне от 500 до 5000 раз.
Читайте также: Все виды ткани оксфорд
Инактивация натриевого канала. В верхней правой части рисунке показано третье состояние натриевого канала. Увеличение потенциала, открывающее активационные ворота, закрывает инактивационные ворота. Однако инактивационные ворота закрываются в течение нескольких десятых долей миллисекунды после открытия активационных ворот. Это значит, что конформационное изменение, приводящее к закрытию инактивационных ворот, — процесс более медленный, чем конформационное изменение, открывающее активационные ворота. В результате через несколько десятых долей миллисекунды после открытия натриевого канала инактивационные ворота закрываются, и ионы натрия не могут более проникать внутрь волокна. С этого момента мембранный потенциал начинает возвращаться к уровню покоя, т.е. начинается процесс реполяризации.
Существует другая важная характеристикая процесса инактивации натриевого канала: инактивационные ворота не открываются повторно до тех пор, пока мембранный потенциал не вернется к значению, равному или близкому к уровню исходного потенциала покоя. В связи с этим повторное открытие натриевых каналов обычно невозможно без предварительной реполяризации нервного волокна.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
7. Фазовые изменения возбудимости клеток при генерации потенциала действия.
Уровень возбудимости клетки зависит от фазы ПД. В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением. В фазу деполяризации ПД, когда открываются все натриевые каналы и ноны натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной не возбудимости или абсолютной рефрактерности, т.е. фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной не возбудимости или относительной рефрактерности. Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно, в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации или супернормальной возбудимости.
В момент следовой гиперполяризации МП выше исходного уровня, т.е. дальше КУД и ее возбудимость снижена. Она наводится в фазе субнормальной возбудимости. Рис. Следует отметить, что явление .аккомодации также связано с изменением проводимости ионных каналов. Если деполяризуюший ток нарастает медленно, то это приводит к частичной инактивации натриевых, и активации калиевых каналов. Поэтому развития ПД не происходит.
Законы раздражения возбудимых тканей: закон силы, закон длительности, закон скорости нарастания раздражения.
Законы устанавливают зависимость ответной реакции ткани от параметров раздражителя. Существуют три закона раздражения возбудимых тканей:
2) закон длительности раздражения;
3) закон градиента раздражения.
Закон силы раздражения устанавливает зависимость ответной реакции от силы раздражителя. Эта зависимость неодинакова для отдельных клеток и для целой ткани. Для одиночных клеток зависимость называется «все или ничего». Характер ответной реакции зависит от достаточной пороговой величины раздражителя.
Закон длительности раздражений. Ответная реакция ткани зависит от длительности раздражения, но осуществляется в определенных пределах и носит прямо пропорциональный характер.
Закон градиента раздражения. Градиент – это крутизна нарастания раздражения. Ответная реакция ткани зависит до определенного предела от градиента раздражения.
Законы действия постоянного тока на возбудимые ткани.
Полярный закон действия тока. При раздражении нерва или мышцы постоянным током возбуждение возникает в момент замыкания постоянного тока только под катодом, а в момент размыкания — только под анодом, причем порог замыкательного удара меньше, чем размыкательного. Прямые измерения показали, что прохождение через нервное или мышечное волокно электрического тока вызывает прежде всего изменение мембранного потенциала под электродами. В области приложения к поверхности ткани анода (+) положительный потенциал на наружной поверхности мембраны возрастает, т.е. в этом участке происходит гиперполяризация мембраны, что не способствует возбуждению, а, наоборот, ему препятствует. В том же участке, где к мембране приложен катод (-), положительный потенциал наружной поверхности снижается, происходит деполяризация, и если она достигает критической величины — в этом месте возникает ПД.
Читайте также: Ткань sense 02 cappuccino
Изменение МП возникают не только непосредственно в точках приложения к нервному волокну катода и анода, но и на некотором расстоянии от них, но величина этих сдвигов убывает по мере удаления от электродов. Изменения МП под электродами носят название электротонических ( соответственно кат-электротон и ан-электротон), а за электродами — периэлектротонических (кат- и ан-периэлектротон).
Увеличение МП под анодом (пассивная гиперполяризация) не сопровождается изменением ионной проницаемости мембраны даже при большой силе приложенного тока. Поэтому при замыкании постоянного тока возбуждение под анодом не возникает. В отличие от этого, уменьшение МП под катодом (пассивная деполяризация) влечет за собой кратковременное повышение проницаемости для Na, что приводит к возбуждению.
Повышение проницаемости мембраны для Na при пороговом раздражении не сразу достигает максимальной величины. В первый момент деполяризация мембраны под катодом приводит к небольшому увеличению натриевой проницаемости и открытию небольшого числа каналов. Когда же под влиянием этого в протоплазму начинают поступать заряженные положительно ионы Na+, то деполяризация мембраны усиливается. Это ведет к открытию других Na-каналов, и, следовательно, к дальнейшей деполяризации, которая, в свою очередь, обуславливает еще большее повышение натриевой проницаемости. Этот круговой процесс, основанный на т.н. положительной обратной связи, получил название регенеративной деполяризации. Возникает она только при снижении Ео до критического уровня (Ек ). Причина повышения натриевой проницаемости при деполяризации связана, вероятно, с удалением Са++ из натриевых ворот при возникновении электро отрицательности (или снижении электро положительности) на наружной стороне мембраны.
Повышенная натриевая проницаемость через десятые доли миллисекунды за счет механизмов натриевой инактивации прекращается.
Скорость, с которой происходит деполяризация мембраны, зависит от силы раздражающего тока. При слабой силе деполяризация развивается медленно, и поэтому для возникновения ПД такой стимул должен иметь большую длительность.
Локальный ответ, который возникает при под пороговых стимулах, так же, как и ПД, обусловлен повышением натриевой проницаемости мембраны. Однако при под пороговом стимуле это повышение недостаточно велико для того, чтобы вызвать процесс регенеративной деполяризации мембраны. Поэтому начавшаяся деполяризация приостанавливается инактивацией и повышением калиевой проницаемости.
Подводя итог изложенному выше, можно следующим образом изобразить цепь событий, развивающихся в нервном или мышечном волокне под катодом раздражающего тока: пассивная деполяризация мембраны —- повышение натриевой проницаемости — усиление потока Na внутрь волокна — активная деполяризация мембраны — локальный ответ — превышение Ек — регенеративная деполяризация — потенциал действия (ПД).
Каков же механизм возникновения возбуждения под анодом при размыкании? В момент включения тока под анодом мембранный потенциал возрастает — происходит гиперполяризация. При этом разница между Ео и Ек растет, и для того, чтобы сдвинуть МП до критического уровня, нужна большая сила. При выключении тока (размыкание) исходный уровень Ео восстанавливается. Казалось бы, в это время нет условий для возникновения возбуждения. Но это справедливо только для того случая, если действие тока продолжалось очень короткое время (менее 100 мсек.). При длительном действии тока начинает меняться сам критический уровень деполяризации — он растет. И, наконец, возникает момент, когда новый Ек становится равным старому уровню Ео. Теперь при выключении тока возникают условия для возбуждения, ибо мембранный потенциал становится равным новому критическому уровню деполяризации. Величина ПД при размыкании всегда больше, чем при замыкании.
Читайте также: Роза из полоски ткани своими руками пошагово
Возбудимость мембраны на разных этапах потенциала действия
При локальном ответе наблюдается увеличение возбудимости мембраны, если раздражитель имеет достаточную силу, длительность, частоту, то локальный ответ переходит в потенциал действия. Первая фаза – деполяризация характеризуется абсолютной рефрактерностью мембраны, она не возбудима в ответ на любые стимулы(сильные или слабые).
Вторая фаза – в период реполяризации наступает фаза относительной рефрактерности, при которой мембрана реагирует только на сильные надпороговые раздражители.
Третья фаза — отрицательная гиппер поляризация в этот период наступает фаза экзальтации, в процессе которой мембрана возбудима для всех раздражителей. Это фаза максимальной возбудимости. Даже подпороговые раздражители могут давать ответную реакцию со стороны мембраны
Четвёртая фаза – положительная гипперполяризация. Мембрана вступает в период суппер нормальной активности, т.е постепенно возвращается к тому исходному знаку заряда, который наблюдался до нанесения раздражения.
Проводимость (проницаемость) мембраны. В период деполяризации мембрана максимально проницаема для ионов натрия. Натрий проникает в клетку через открытые активационные и инактивационные ворота натриевого канала.
Ионные каналы для калия имеют одну створку. В фазу деполяризации она закрыта, поэтому калий не может покинуть клетку следовательно проницаемость для калия крайне низкая.
В период реполяризации снижается проницаемость для ионов натрия, поскольку закрываются внутренние инактивационные ворота и натрий не может проникнуть внутрь клетки. Ионные каналы для калия открываются, проницаемость увеличивается и калий свободно выходит за пределы клетки. В период гиппер поляризации отрицательной. Работают хлорные каналы, засчет чего усилена проницаемость для хлора и нулевая проницаемость для натрия и низкая проницаемость для калия. Калий может только выходить из клетки.
Положительная гипперполяризация. Проницаемость для хлора резко снижается. Происходит проникновение ионов натрия через калий натриевый насос.
Локальный ответ – это процесс, который развивается на мембране при действии более слабого раздражителя или на начальном этапе потенциала действия. В отличии от потенциала действия, локальный ответ распространяется с затуханием или с декрементом на один, два волокна, в то время как потенциал действия распространяется по всему волокну. Величина локального ответа увеличивается в связи с увеличением силы раздражителя, т.е подчиняется закону силы. Чем сильнее раздражитель, тем больше величина локального ответа. Потенциал действия подчинятся закону «Всё или ничего», т.е раздражителю достаточно достигнуть пороговой величины и возникает потенциал действия (это всё). Если раздражитель не достиг пороговой величины, потенциал действия не происходит (это ничего). Локальный ответ обладает способностью к суммации, в отличии от потенциала действия. Его амплитуда составляет от 10 до 40 милливольт. У потенциала действия 100-130 милливольт. Возбудимость мембраны при локальном ответе увеличивается. При потенциале действия — снижается.
Вопросы семинара: (презентация)
Строение мембраны нервных клеток
Функции и свойства мембран нервных клеток. Теории Бернштейна.
Транспортная функция мембраны.
Мембранный потенциал и причины его возникновения.
Потенциал действия и причины его возникновения.
Проводимость мембраны или проницаемость на разных этапах потенциала действия.
Возбудимость мембраны на разных этапах действия.