
Во внутриклеточной жидкости возбудимых тканей высока концентрация ионов
Плазматическая мембрана всех клеток, в том числе и нейронов, представлена двойным слоем (бислоем) фосфолипидов, в котором «головки» фосфолипидов обращены к водной среде внеклеточных и внутриклеточных пространств, а парные «хвосты» погружены внутрь и образуют липидную мембрану. Фосфатный слой растворим в воде (гидрофильный, или полярный), а двойной липидный слой — нерастворим (гидрофобный, или неполярный).
Внеклеточная и внутриклеточная жидкости представляют собой водно-солевые растворы, в которых множество растворимых молекул диссоциируют на положительно и отрицательно заряженные атомы,— ионы. Ионы и молекулы водных растворов находятся в состоянии постоянного возбуждения и осуществляют процесс диффузии—движения из областей высокой концентрации в область меньшей концентрации. Диффузия ионов происходит не только по градиенту концентрации, но и под влиянием электрических градиентов. Положительно заряженные ионы, например ионы натрия (Na + ) и калия (К + ), называют катионами, поскольку в электрическом поле они движутся к катоду.
Отрицательно заряженные ионы, например ионы хлора (Cl — ), называют анионами, так как они движутся к аноду. Одноименные заряды (например, Na + и К + ) отталкиваются, а разноименные (например, Na + и Cl — ) — притягиваются.
Клеточную мембрану можно рассматривать в качестве электрического конденсатора, поскольку она состоит из двух противоположно заряженных слоев, между которыми расположен изолирующий липидный слой. Электрический потенциал тканевой жидкости на плазматической мембране стремится к нулю (0 мВ) за счет нейтрализующих влияний анионов Cl — на катионы Na + (и другие катионы), а в цитозоле — за счет влияний белков-анионов на катионы К + .
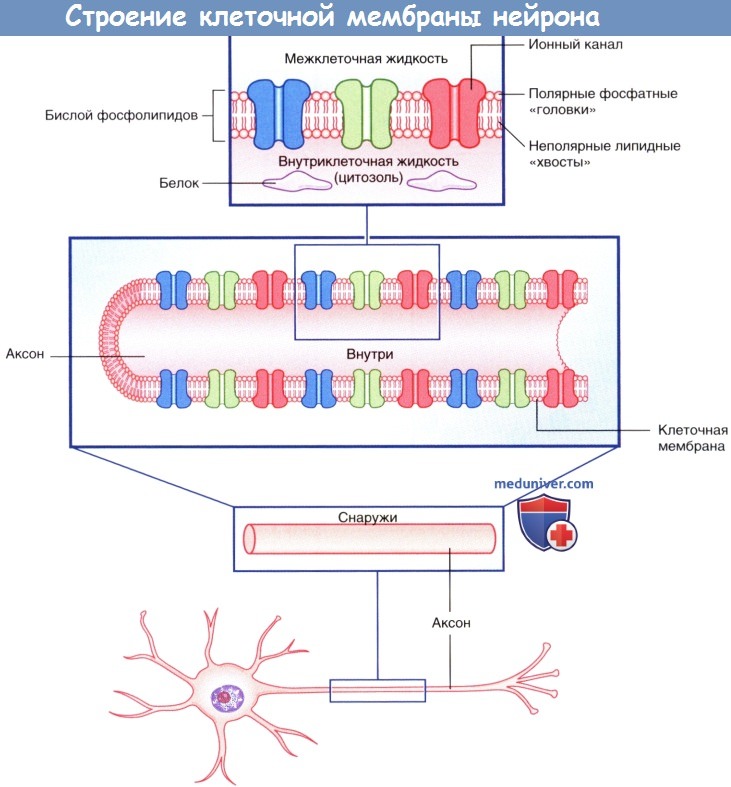
Строение клеточной мембраны нейрона.
Изображены мембранные белки, образующие ионные каналы.
а) Ионные каналы. Ионные каналы представляют собой трансмембранные белки, в центре которых находятся отверстия, по которым осуществляется транспорт ионов через клеточную мембрану. Большинство ионных каналов селективно пропускает определенные ионы, например, Na + , К + , Cl — . Выделяют несколько видов ионных каналов, из которых наиболее значимы первые три типа из следующих:
• Неуправляемые (независимые) ионные каналы постоянно находятся в открытом состоянии и осуществляют трансмембранный перенос ионов, создавая потенциал покоя на мембранах нейронов.
• Потенциалозависимые (потенциал-управляемые) ионные каналы содержат потенциалчувствительную цепь аминокислот, регулирующую открытие и закрытие отверстий ионных каналов в зависимости от изменений мембранного потенциала. Потенциал-управляемые ионные каналы играют важную роль в формировании потенциала действия.
• Энергозависимые транспортеры — ионные обменники (насосы) — обеспечивают постоянство концентраций ионов. Натрий-калиевый насос обеспечивает поддержание потенциала покоя.
• Хемоуправляемые (медиаторозависимые) ионные каналы нервной системы обеспечивают временное преобразование мембранного потенциала. В основном эти каналы представлены на постсинаптических мембранах. Активация хемоуправляемых ионных каналов может осуществляться молекулами медиаторов напрямую или опосредованно.
• Механочувствительные каналы активируются под действием физических раздражителей, вызывая деполяризацию мембраны и образование потенциалов действия, что обеспечивает восприятие определенных сигналов нервной системой. Каждый рецептор осуществляет преобразование специфических сигналов, например изменения длины или сократимости мышечных волокон, температурной и тактильной чувствительности кожи, хемочувствительности полости носа и рта или электромагнитных импульсов сетчатки.
На рисунке ниже изображены три неуправляемых ионных канала, обеспечивающих развитие потенциала покоя.
Если бы пассивная диффузия различных ионов через соответствующие им каналы осуществлялась свободно в одинаковом объеме, то не существовало бы разницы зарядов внутри и снаружи мембраны. В действительности проницаемость плазматической мембраны для ионов К + и Cl — намного выше, чем для ионов Na + , поскольку количество ионных каналов различается: калиевые каналы наиболее многочисленны, количество хлорных каналов вдвое меньше, а натриевые каналы встречают реже всего.
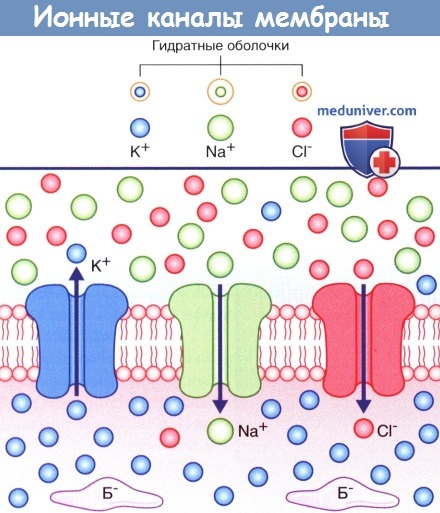
В состоянии покоя ионы Na + и Cl — сконцентрированы на внешней стороне плазматической мембраны за счет взаимного притяжения и образования толстой гидратной оболочки вокруг ионов Na + .
Ионы К + сконцентрированы на внутренней стороне плазматической мембраны за счет притяжения к белкам-анионам (Б — ).
Стрелками показано направление ионных градиентов концентрации.
б) Мембранный потенциал покоя. Мембранный потенциал невозбужденного нейрона (находящегося в состоянии покоя) формируется за счет разницы концентраций ионов Na + и К + во внеклеточной жидкости и цитозоле. Концентрация ионов К + в цитозоле выше, чем во внеклеточной жидкости в 20 раз, в то время как концентрация ионов Na + и Cl — во внеклеточной жидкости выше, чем в цитозоле в 10 и 3,8 раз, соответственно. Это обусловлено тем, что ионы К + под действием химических движущих сил переносятся из клетки в межклеточную жидкость, а ионы Na+ и Cl — — в обратном направлении.
На рисунке ниже показан вольтметр, электроды которого погружены в окружающую аксон внеклеточную жидкость. Один из электродов находится в стеклянной пипетке с тонким кончиком. В левой части рисунка, где оба электрода погружены во внеклеточную жидкость, разница электрического напряжения отсутствует, и вольтметр регистрирует нулевой показатель. В правой части рисунка пипетка проникает через плазматическую мембрану аксона в цитозоль, что позволяет оценить электрический заряд внутриклеточной жидкости. В данном случае разность электрических потенциалов составляет -70 мВ, однако у разных нейронов она может варьировать от -60 до -80 мВ.
Читайте также: Ткань crown 230 свойства
Этот показатель отражает мембранный потенциал покоя, т.е. потенциал мембраны аксона, который не проводит электрические импульсы.
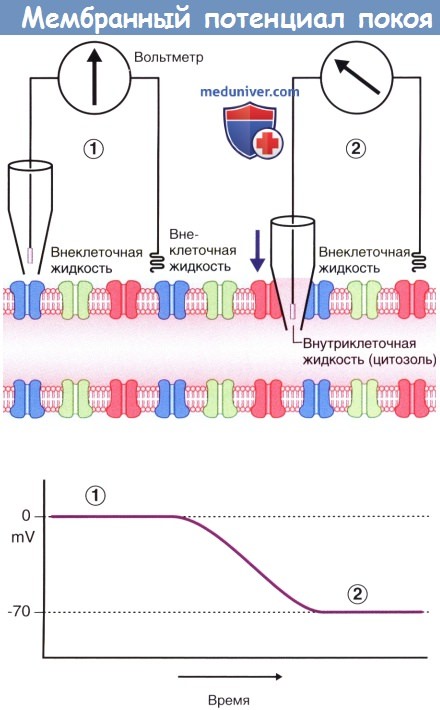
Мембранный потенциал покоя.
(1) Оба электрода вольтметра погружены во внеклеточную жидкость, окружающую аксон. Конец левого электрода находится в стеклянной пипетке.
Разница электрических потенциалов не регистрируется; на графике отмечены нулевые показатели.
(2) При опускании пипетки (направление указано стрелкой) производят пункцию плазматической мембраны для забора внутриклеточной жидкости.
Регистрируется разница электрических потенциалов величиной -70 мВ.
в) Проницаемость мембраны в состоянии покоя: 1. Ионы К + . На уровне ионного канала перемещение ионов К + осуществляется под электрическими влияниями притягивающего действия белков-анионов на внутренней стороне мембраны и отталкивающего действия ионов Na + на ее наружной стороне; при отсутствии этих факторов концентрация ионов К + на разных сторонах плазматической мембраны была бы одинаковой.
Концентрационный градиент на внешней стороне плазматической мембраны и электрический градиент потенциала на внутренней ее стороне уравновешиваются, когда мембранный потенциал достигает определенной величины, которую называют равновесным потенциалом для ионов К + (Ek). Его величина может быть рассчитана из уравнения Нернста, которое позволяет представить электрический градиент потенциала иона через его концентрационный градиент на основании законов термодинамики:
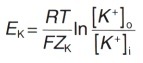
где Ek — равновесный потенциал для ионов К + (мВ)
R — газовая постоянная (8,31 Дж/(моль/°К)
Т — абсолютная температура в градусах Кельвина (310 К = 37 °С) F — число Фарадея (96500 Кл/моль)
Zk — валентность ионов К + (+1)
[К + ]0 — концентрация ионов K + с наружной стороны мембраны
[К + ]i — концентрация ионов К + с внутренней стороны мембраны.
Переведем натуральный логарифм в десятичный и рассчитаем значение показателя дроби:
Рассчитаем эти же значения для ионов Na + и Cl — :

Для расчета мембранного потенциала покоя применяют уравнение Гольдмана, учитывающее проницаемость мембраны для трех основных ионов и их приблизительное количество.
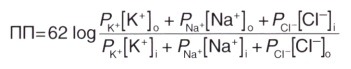
62 — RT/Fx2,3 (постоянная для перевода натурального логарифма в десятичный)
Р — константа проницаемости мембраны для каждого из ионов (соответствует приблизительному количеству ионных каналов) о, i—концентрации ионов на внешней и внутренней сторонах мембраны, соответственно; концентрация отрицательно заряженных ионов Cl — внутри клетки расположена в числителе, а снаружи — в знаменателе дроби, поскольку log (X/Y) = log (Y/X).
Квадратными скобками обозначены концентрации ионов.
Уравнение Гольдмана сходно с уравнением Нернста, однако в нем перемножаются проницаемость мембраны для каждого иона и его концентрация; это уравнение применяют для установления потенциала «реверсии» через плазматическую мембрану с учетом ее проницаемости для разных ионов. Влияние ионов С1″ на потенциал покоя имеет второстепенное значение, поскольку равновесный потенциал и потенциал покоя ионов Cl — приблизительно равны. Значение потенциала покоя в среднем составляет -70 мВ, и обеспечивается за счет суммарного влияния ионов К + и Na + .
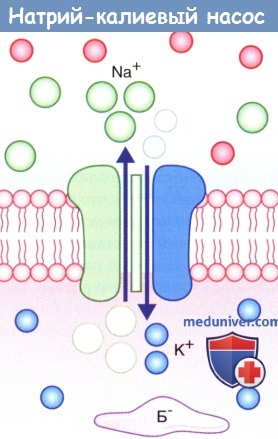
Натрий-калиевый насос.
На рисунке показан одновременный перенос трех ионов Na + из клетки в обмен на два иона К + в клетку.
Б — — белок-анион.
2. Натрий-калиевый насос. Под влиянием градиента концентрации происходит непрерывное перемещение ионов Na + внутрь клетки, а ионов К + — наружу, что нарушает постоянство потенциала покоя. Поддержание этого равновесия обеспечивает натрий-калиевый насос, корректирующий пассивное движение ионов. Натрий-калиевый насос представляет собой канал, способный перемещать ионы Na + наружу, а ионы К + — внутрь клетки. Во время работы насоса происходит перенос трех ионов Na + из клетки на каждые два иона К + в клетку.
Перенос обоих катионов осуществляется против градиента концентрации за счет энергии превращения АТФ в АДФ ферментом АТФазой. Активация этого фермента происходит при повышении концентрации ионов Na + в цитозоле.
Аксональная дегенерация при рассеянном склерозе возникает вследствие нарушения работы натрий-калиевого насоса в демиелинизированной аксолемме, что вызывает увеличение концентрации ионов Na + , которое в свою очередь приводит к повышению высвобождения ионов Са 2+ из внутриклеточного пула.
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
Физиология и свойства возбудимых тканей.
Клетка – это элементарная живая система, состоящая из ядра и цитоплазмы, лежащая в основе развития, строения и функции всех живых и растительных организмов.
В каждой клетке есть «органы»:
2. Цитоплазма и оболочка, которые образованы живым веществом клетки – протоплазмой.
Читайте также: Нейлон ткань для верхней одежды
Кроме «органов» в цитоплазме клетки имеются образования, выполняющие определенные функции — это органоиды илиорганеллы.
Некоторые органоиды есть во всех видах клеток, они называются органоидами общего порядка. К ним относятся:
Все эти образования выполняют определенную функцию.
Ядро – регулирует жизнедеятельность клетки, осуществляет передачу генетической информации, синтез белка и рецепцию биологически активных веществ.
Цитоплазма участвует в процессах метаболизма и поддержании постоянства внутренней среды клетки.
Эндоплазматическая сеть является главным депо ионов Са
Рибосомы синтезируют белки
Митохондрии участвуют в генерации и аккумуляции энергии
Аппарат Гольджи (пластинчатый комплекс) участвует в секреции биологически активных веществ.
Лизосомы осуществляют переваривание поглощенных клетками питательных веществ.
Кроме органоидов общего порядка есть органоиды, которые встречаются в клетках определенного вида. Такие органоиды называются специальными. К ним относятся:
Миофибриллы (мышечная ткань)
Нейрофибриллы (нервная ткань)
Реснички и жгутики (как остатки органоидов движения одноклеточных, в эпителиальных тканях)
Ультраструктура биологических мембран.
Структурной основой биологической мембраны является двойной слой фосфолипидов, в который встроены мембранные белки.
Белки, пронизывающие насквозь фосфолипидный слой, называются внутренними мембранными белками, или белковыми каналами, или порами.
В функциональном отношении мембранные белки делятся на 4 класса: «насосы», каналы, рецепторы и ферменты.
«Насосы» расходуют метаболическую энергию АТФ для перемещения ионов и молекул против концентрационных и электрохимических градиентов и поддерживают необходимые концентрации этих молекул в клетке.
Ионоселективные каналы представляют собой пути переноса заряженных молекул и ионов. Через каналы в клетку проникают и лекарственные вещества.
Рецепторы мембран представлены белковыми молекулами, которые «узнают» то или иное биологически активное вещество, контактируют с ним и передают в клетку информацию о характере биохимических взаимодействий.
Белки-ферменты, обладающие высокой каталитической активностью, облегчают протекание биохимических реакций как внутри мембраны, так и у ее поверхности.
Функции биологических мембран.
— Пограничная функция. Мембрана отграничивает цитоплазму от межклеточной жидкости, а большинство внутриклеточных структур: митохондрии, ядро, эндоплазматическую сеть – от цитоплазмы.
— Биотрансформирующая функция. Любое вещество, проходя через мембрану, вступает с ней в сложное взаимодействие и претерпевает ряд биохимических превращений. В результате биотрансформации лекарственное вещество, как правило, переходит в форму, легко усвояемую клеткой.
— Транспортная функция. Перенос веществ через биологические мембраны связан с процессами метаболизма, поддержанием постоянства внутренней среды клетки, возбуждением и проведением нервного импульса. Существует два основных типа переноса: пассивный (фильтрация, диффузия, облегченная диффузия, осмос) и активный (работа мембранных белковых «насосов»)
Пассивный транспорт. Фильтрация осуществляется через мембранные белковые каналы – поры, зависит от разности давлений снаружи и внутри клетки и проницаемости мембраны для жидкости и низкомолекулярных веществ. Диаметр пор чрезвычайно мал, поэтому фильтруются только низкомолекулярные вещества, вода и некоторые ионы.
Диффузия — пассивное передвижение молекул или ионов по градиенту концентрации (из области высокой концентрации в область низкой). Осмос представляет собой частный случай диффузии растворителя через полупроницаемую мембрану, не пропускающую растворенные вещества.
Пассивный транспорт не требует затрат энергии.
Активный транспорт. Это универсальный для всех видов мембран перенос веществ против концентрационных или электрохимических градиентов (из области низкой концентрации в область высокой). При помощи активного транспорта переносятся гидрофильные полимерные молекулы, неорганические ионы (Na, Ca, K) , водород, сахара, аминокислоты, витамины, гормоны и лекарственные вещества. Активный транспорт осуществляется с обязательной затратой энергии, образующейся при расщеплении (окислительное фосфорилирование) аденозинтрифосфорной кислоты (АТФ).
Разновидностью активного транспорта, связанной с деятельностью самой клетки, является микровезикулярный транспорт (пиноцитоз, экзоцитоз и фагоцитоз). При пиноцитозе происходит активное поглощение клеткой жидкости из окружающей среды с формированием пузырьков и последующим переносом их через цитоплазму. Процесс слияния пузырьков с мембраной клетки и выделение клеткой вещества в виде секреторных гранул или вакуолей называется экзоцитозом. Явление фагоцитоза заключается в способности клеток активно захватывать и поглощать микроорганизмы, разрушенные клетки и инородные частицы.
— Рецепторная функция. Биологические мембраны имеют большое количество рецепторов – участков, молекулярная структура которых характеризуется избирательным сродством к определенным физиологически активным веществам: гормонам, медиаторам, антигенам.
— Образование межклеточных контактов.
— Генерация биоэлектрических потенциалов. В ходе эволюции у железистого эпителия, мышечной и нервной тканей появилось свойство возбудимости – способность реагировать на воздействие окружающей среды возбуждением. Внешним проявлением возбуждения является возникновение биоэлектрического потенциала.
Все ткани организма могут находиться в двух состояниях:
состоянии относительного физиологического покоя;
Наблюдается при раздражении ткани. Существует 2 вида активного состояния тканей: возбуждение и торможение.Возбуждение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся повышением функций ткани. Возбуждение характеризуется двумя группами признаков:неспецифическими и специфическими.
Читайте также: Белые ткани для свадебных платьев
Неспецифические признаки возникают у всех возбудимых тканей вне зависимости от их строения:
изменение проницаемости клеточных мембран
изменение заряда клеточных мембран,
повышение потребления кислорода
усиление обменных процессов
Специфические признаки различаются у различных тканей:
мышечная ткань – сокращение
железистая ткань – выделение секрета
нервная ткань – генерация нервного импульса.
Процесс возбуждения связан с наличием в мембране электрически (для ионов кальция и хлора) и химически (для ионов натрия и калия) управляемых каналов, которые могут открываться в ответ на соответствующее раздражение клетки.
Ионоселективные каналы. Для каждого из переносимых через мембрану вида ионов существуют самостоятельные транспортные системы – ионные каналы (натриевые, калиевые, кальциевые, каналы для хлора и т.д.). Ионный каналсостоит из поры, воротного механизма, сенсора (индикатора) напряжения ионов в самой мембране и селективного фильтра.
Пора представляет собой молекулярное динамическое образование, которое может находиться в открытом и закрытом состоянии. Образована пора «транспортным» ферментом – белком с высокой каталитической активностью, который способен переносить ионы через мембрану со скоростью в 200 раз превышающей скорость простой диффузии.
Воротный механизм (ворота канала) расположен на внутренней стороне мембраны и представлен белковыми молекулами, способными к конформации (изменение пространственной конфигурации молекул). В тысячные доли секунды он открывает (активирует) и закрывает (инактивирует) канал и таким образом регулирует скорость передвижения ионов по нему и поступление их в цитоплазму.
Сенсор напряжения ионов в мембране представлен белковой молекулой, расположенной в самой мембране и способной реагировать на изменение мембранного потенциала.
Селективный фильтр находится в самом узком месте канала. Он определяет однонаправленное движение ионов через пору и ее избирательную проницаемость.
В развитии возбуждения выделяют 4 этапа:
1) предшествующее возбуждению состояние покоя (статическая поляризация);
Статическая поляризация – наличие постоянной разности потенциалов между наружной и внутренней поверхностями клеточной мембраны. В состоянии покоя наружная поверхность клетки всегда электроположительна по отношению к внутренней, т.е. поляризована. Эта разность потенциалов, равная
60 мВ, называется потенциалом покоя, илимембранным потенциалом (МП). В образовании потенциала принимают участие 4 вида ионов: катионы натрия (положительный заряд), катионы калия (положительный заряд), анионы хлора (отрицательный заряд), анионы органических соединений (отрицательный заряд). Во внеклеточной жидкости высока концентрация ионов натрия и хлора, во внутриклеточной жидкости – ионов калия и органических соединений. В состоянии относительного физиологического покоя клеточная мембрана хорошо проницаема для катионов калия, чуть хуже для анионов хлора, практически непроницаема для катионов натрия и совершенно непроницаема для анионов органических соединений.
В покое ионы калия без затрат энергии выходят в область меньшей концентрации (на наружную поверхность клеточной мембраны), неся с собой положительный заряд. Ионы хлора проникают внутрь клетки, неся отрицательный заряд. Ионы натрия продолжают оставаться на наружной поверхности мембраны, еще больше усиливая положительный заряд.
Деполяризация – сдвиг МП в сторону его уменьшения. Под действием раздражения открываются «быстрые» натриевые каналы, вследствие чего ионы Na лавинообразно поступают в клетку. Переход положительно заряженных ионов в клетку вызывает уменьшение положительного заряда на ее наружной поверхности и увеличение его в цитоплазме. В результате этого сокращается трансмембранная разность потенциалов, значение МП падает до 0, а затем по мере дальнейшего поступления Na в клетку происходят перезарядка мембраны и инверсия ее заряда (поверхность становится электроотрицательной по отношению к цитоплазме) – возникает потенциал действия (ПД). Электрографическим проявлением деполяризации является спайк, или пиковый потенциал.
Во время деполяризации, когда переносимый ионами Na положительный заряд достигает некоторого порогового значения, в сенсоре напряжения ионных каналов возникает ток смещения, который «захлопывает» ворота и «запирает» (инактивирует) канал, прекращая тем самым дальнейшее поступление Na в цитоплазму. Канал «закрыт» (инактивирован) вплоть до восстановления исходного уровня МП.
Реполяризация – восстановление исходного уровня МП. При этом ионы натрия перестают проникать в клетку, проницаемость мембраны для калия увеличивается, и он достаточно быстро выходит из нее. В результате заряд клеточной мембраны приближается к исходному. Электрографическим проявлением реполяризации является отрицательныйследовой потенциал.
Гиперполяризация – увеличение уровня МП. Вслед за восстановлением исходного значения МП (реполяризация) происходит его кратковременное увеличение по сравнению с уровнем покоя, обусловленное повышением проницаемости калиевых каналов и каналов для Cl . В связи с этим поверхность мембраны приобретает избыточный по сравнению с нормой положительный заряд, а уровень МП становится несколько выше исходного. Электрографическим проявлением гиперполяризации является положительный следовой потенциал. На этом заканчивается одиночный цикл возбуждения.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
