
Клетки миокарда обладают возбудимостью, но им не присуща автоматия. В период диастолы мембранный потенциал покоя этих клеток стабилен, и его величина выше (80—90 мВ), чем в клетках водителей ритма. Потенциал действия в этих клетках возникает под влиянием возбуждения клеток водителей ритма, которое достигает кардиомиоцитов, вызывая деполяризацию их мембран.
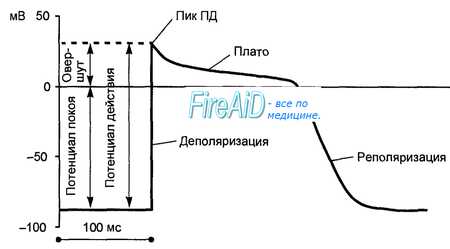
Рис. 9.8. Потенциал действия клетки рабочего миокарда. Быстрое развитие деполяризации и продолжительная реполяризация. Замедленная реполяри-зация (плато) переходит в быструю реполяризацию.
Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации, начальной быстрой реполяризации, переходящей в фазу медленной реполяризации (фаза плато), и фазы быстрой конечной реполяризации (рис. 9.8). Фаза быстрой деполяризации создается резким повышением проницаемости мембраны для ионов натрия, что приводит к возникновению быстрого входящего натриевого тока. Последний, однако, при достижении мембранного потенциала 30—40 мВ инактивируется и в последующем, вплоть до инверсии потенциала (около +30 мВ) и в фазу «плато», ведущее значение имеют кальциевые ионные токи. Деполяризация мембраны вызывает активацию кальциевых каналов, в результате чего возникает дополнительный деполяризирующий входящий кальциевый ток.
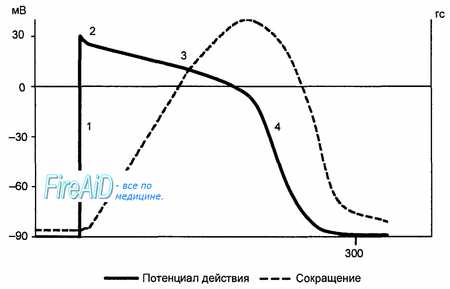
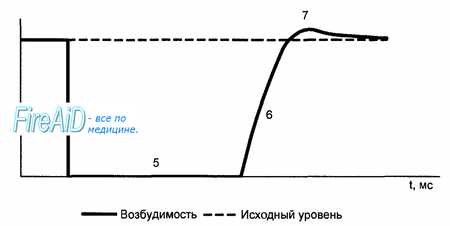 Рис. 9.9. Сопоставление потенциала действия и сокращения миокарда с фазами изменения возбудимости. 1 — фаза деполяризации; 2 — фаза начальной быстрой реполяризации; 3 — фаза медленной реполяризации (фаза плато); 4 — фаза конечной быстрой реполяризации; 5 — фаза абсолютной рефрактерности; 6 — фаза относительной рефрактерности; 7 — фаза супернормальной возбудимости. Рефрактерность миокарда практически совпадает не только с возбуждением, но и с периодом сокращения.
Рис. 9.9. Сопоставление потенциала действия и сокращения миокарда с фазами изменения возбудимости. 1 — фаза деполяризации; 2 — фаза начальной быстрой реполяризации; 3 — фаза медленной реполяризации (фаза плато); 4 — фаза конечной быстрой реполяризации; 5 — фаза абсолютной рефрактерности; 6 — фаза относительной рефрактерности; 7 — фаза супернормальной возбудимости. Рефрактерность миокарда практически совпадает не только с возбуждением, но и с периодом сокращения.
Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя. Длительность потенциала действия кардиомиоцитов составляет 300—400 мс, что соответствует длительности сокращения миокарда (рис. 9.9).
Возбудимость сердца функция ткани

Основы электрофизиологии сердца
Основными функциями клеток сердца являются: функция автоматизма, функция возбудимости, функция рефрактерности, функция проводимости, и функция сократимости.
Повторю, автоматизм – способность клеток автоматически вырабатывать электрические импульсы.
Клетки, автоматически вырабатывающие электрические импульсы, образуют центры автоматизма.
Выделяют центры автоматизма I, II, III порядка.
Центром автоматизма I порядка являются Р-клетки синусового узла, который находится в верхнезадней части правого предсердия — между устьями полых вен.
В нем в норме возникают импульсы частотой у взрослых — 60-80 в 1 минуту и до 140 у маленьких детей.
Центром автоматизма II порядка являются Р-клетки правого и левого предсердия, Р-клетки АВ-соединения (в NH-зоне). Частота импульсов ц центрах II порядка меньшая, обычно 40-60 в 1 минуту.
Центрами автоматизма III порядка являются клетки пучка Гиса и волокон Пуркинье. Частота импульсов в них — 20-40 в 1 минуту.
В норме водитель ритма – синусовый узел (номотопный центр автоматизма)\
Другие центры автоматизма являются ненормальными — гетеротопными или эктопическими.
Очень важно запомнить, что в норме автоматизм эктопических центров подавляется синусовым узлом и нижележайшие центры являются запасными, страхующими сердце от катастрофы. Автоматизм эктопических центров проявляется при угнетении автоматизма синусового узла или при повышении автоматизма эктопических центров — при нарушении проведения электрических импульсов от синусового узла по типу блокады, при нарушении проведения электрических импульсов от синусового по типу re-entrу и др. Обычные клетки миокарда желудочков не обладают способностьч автоматизма. Надо также знать, что синусовый узел и АВ-узел имеют симпатическую и парасимпатическую иннервацию и, следовательно подвержен всевозможным экстракардиальным влияниям. Возбудимость — это способность сердца возбуждаться под влиянием импульсов. Функцией возбудимости обладают клетки как проводящей системы сердца, так и сократительного миокарда. Возбуждение сердечной мышцы сопровождается возникновением трансмембранного потенциала действия (ТМПД) и в конечном счете — электрического тока.
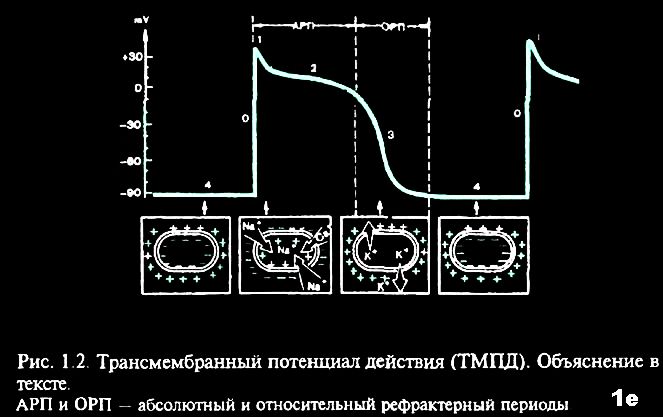
В разные фазы ТМПД возбудимость мышечного волокна при поступлении нового импульса различна. В начале ТМПД клетки полностью невозбудимы, или рефрактерны к дополнительному электрическому импульсу (1,2). Это так называемый абсолютный рефрактерный период миокардиального волокна, когда клетка вообще неспособна отвечать новой активацией на какой-либо дополнительный электрический стимул В конце ТМПД имеет место относительный рефрактерный период, во время которого нанесение очень сильного дополнительного стимула может привести к возникновению нового повторного возбуждения клетки, тогда как слабый импульс остается без ответа (3). Во время диастолы полностью восстанавливается возбудимость миокардиального волокна, а его рефрактерность отсутствует (4). Значение активных сил в формировании мембранного потенциала. Перемещение ионов происходит путем диффузии. Активный транспорт осуществляется за счет Nа+-К+ насоса (Р. Дин — 1941 г.). Nа+-К+ насос осуществляет движение ионов против градиента концентрации (К+ внутрь, Nа+ — наружу). Для работы насоса требуется энергия, которая образуется при расщеплении АТФ под влиянием АТФ-азы, которая активизируется при изменении концентрации К+ и Nа+, что происходит постоянно, поэтому Nа+-К+ насос работает постоянно. Согласно Дину движение ионов осуществляется за счет молекул переносчиков (белки внутри клеточных мембран). После выполнения функции Х-белок (переносчик ионов К+), благодаря энергии АТФ, меняет свою структуру и превращается в У-белок (переносчик ионов Nа+). Nа+-К+ насоса неодинакова при различных состояниях. В состоянии покоя на 3 иона Nа+ приходится 2 иона К+. При изменении состояния клетки меняется активность Nа+-К+ насоса.
Читайте также: Плед из ализе пуффи с подкладкой из ткани
Итак, в состоянии покоя за счет выхода ионов К+ из клетки наружная поверхность клетки заряжена положительно, а внутренняя — отрицательно (по отношению к наружной поверхности). Это состояние называется поляризация; мембранный потенциал является равновесным калиевым потенциалом; в возникновении мембранного потенциала участвуют другие ионы и активные силы. Механизм формирования потенциала действия.
Потенциал действия возникает в ткани под влиянием порогового и сверхпорогового раздражителей и является импульсивным возбуждением. Потенциал действия можно так же, как и мембранный потенциал, зарегистрировать трансмембранным способом. Под влиянием пороговых раздражителей изменяется проницаемость клеточной мембраны — повышается для всех потенциалобразующих ионов, но больше всего для ионов Nа+ (в 500 раз). Ионы натрия перемещаются внутрь клетки. Движение ионов натрия внутрь клетки превышает выход ионов К+ из клетки. В результате происходит изменение заряда клеточной мембраны на противоположный, затем происходит постепенное восстановление исходного заряда мембраны.  Компоненты потенциала действия и механизм их возникновения.
Компоненты потенциала действия и механизм их возникновения.
При трансмембранном способе регистрации возникает потенциал действия, состоящий из 3-х основных компонентов:
1 компонент: местный (локальный ответ);
2 компонент: пик (спайк);
3 компонент: следовые потенциалы (отрицательный и положительный).
Спайк (пик) — самая постоянная часть. Он состоит из восходящего колена (фаза деполяризации) и нисходящего колена (реполяризация). Остальные компоненты изменчивы и могут отсутствовать. Местный (локальный) ответ возникает и продолжается до тех пор, пока раздражитель не достигнет пороговой величины. Если раздражитель (его сила) меньше 50-75 % пороговой величины проницаемость мембраны изменяется незначительно и равновесно для всех ионов (неспецифично). После достижения силы раздражителя 50-75 % начинает преобладать натриевая проницаемость, т. к. натриевые каналы освобождаются от ионов Са2+. Происходит снижение мембранного потенциала при достижении пороговой величины разность потенциалов достигает критического уровня деполяризации. Критический уровень деполяризации (Ек) — это та разность потенциалов, которая должна быть достигнута, чтобы местные изменения перешли в пик потенциала действия. Ек — пороговая величина, при которой местные изменения переходят в распространенные. Ек величина практически постоянная и равна — 40 — -50 мВ. Разность между мембранным потенциалом и пороговой величиной характеризует порог раздражения и отражает возбудимость ткани. Пик потенциала действия состоит из следующих фаз.
Фаза деполяризации возникает в результате лавинообразного движения Nа+ внутрь клетки. Этому способствуют две причины: открываются потенциалзависимые Nа+-каналы. В этом случае происходит деполяризация по типу процесса с положительной обратной связью (самоподкрепляющийся процесс).
Освобождение натриевых каналов от Са2+.
Заряд клеточной мембраны сначала снижается до 0 (это собственно деполяризация), а затем меняется на противоположный (инверсия или овершут). Для характеристики фазы деполяризации вводится понятие реверсии — это та разность потенциалов, на которую потенциал действия превышает потенциал покоя.
Р=(потенциал действия) — (мембранный потенциал) 20-30 = 50-60 мВ.
Р (реверсия) — это то количество мВ на которое произошла перезарядка мембраны. Фаза деполяризации продолжается до достижения электрохимического равновесия по Nа+. Затем наступает следующая фаза. Амплитуда потенциала действия не зависит от силы раздражителя. Она зависит от концентрации Nа+ (как снаружи, так и внутри клетки), от количества натриевых каналов, особенностей натриевой проницаемости. Фаза реполяризации характеризуется:
снижением проницаемости клеточной мембраны для Nа+ (Nа-инактивация). Натрий накапливается на наружной поверхности клеточной мембраны;
возрастанием проницаемости мембраны для К+, в результате повышается выход К+ из клетки с увеличением положительного заряда на мембране;
изменением активности Nа+-К+ насоса.  Реполяризация — это процесс восстановления заряда мембраны. Но полного восстановления нет, т. к. возникают следовые потенциалы.
Реполяризация — это процесс восстановления заряда мембраны. Но полного восстановления нет, т. к. возникают следовые потенциалы.
Следовые потенциалы делятся на:
Отрицательный следовой потенциал — замедление реполяризации клеточной мембраны. Это результат проникновения внутрь клетки какого-то количества Nа+, таким образом, отрицательный следовой потенциал — это следовая деполяризация.
Положительный следовой потенциал — увеличение разности потенциалов. Это результат повышенного выхода ионов К+ из клетки. Положительный следовой потенциал — это следовая гиперполяризация. Как только калиевая проницаемость возвращается к исходному уровню — регистрируется мембранный потенциал. Проводимость — способность клеток проводить электрические импульсы
электрические импульсы проводят клетки проводящей системы сердца и кардиомиоциты. В норме проводящая система сердца для проведения электрических импульсов от синусового узла включает кардиомиоциты предсердий, АВ-узел, пучок Гиса, правая и левая ножки пучка Гиса, волокна Пуркинье.
Скорость проведения импульсов в предсердиях 1м/с, АВ-узле 0,2м/с, пучке Гиса 1м/с, в ножках и волокнах Пуркинье 3-4м/с.
В норме такая проводящая система определяет последовательность проведения возбуждения в сердце синусового узла. От синусового узла электрические импульсы проводятся на кардиомиоциты предсердий.
В предсердиях электрические импульсы проводятся от правого предсердия на левое предсердие по пучку Бахмана, причем все предсердия возбуждаются за 0,1с.
По кардиомиоцитам предсердий электрические импульсы проводятся на АВ-узел.
По АВ-узлу электрические импульсы проводятся с низкой скоростью – происходит задержка проведения. Эта задержка физиологична – в итоге, систола желудочков наступает после систолы предсердий.
От АВ-узла электрические импульсы проводятся на пучок Гиса, ножки пучка Гиса, волокна Пуркинье и далее на кардиомиоциты желудочков.
В желудочках электрические импульсы распространяются от средней части межжелудочковой перегородки на верхушку правого желудочка, затем на верхушку левого желудочка, затем на базальную часть желудочков и перегородки
Все желудочки возбуждаются за 0,1с, причем распространяется от эндокарда к эпикарду.
Читайте также: Режущий инструмент для обработки ткани 7 букв
Мышечные ткани

Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей). Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.
Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры. Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.  Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро. Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим). Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности. Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.
Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро. Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим). Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности. Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.  Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань

Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок. В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы). Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400). Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани. Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).  Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез). 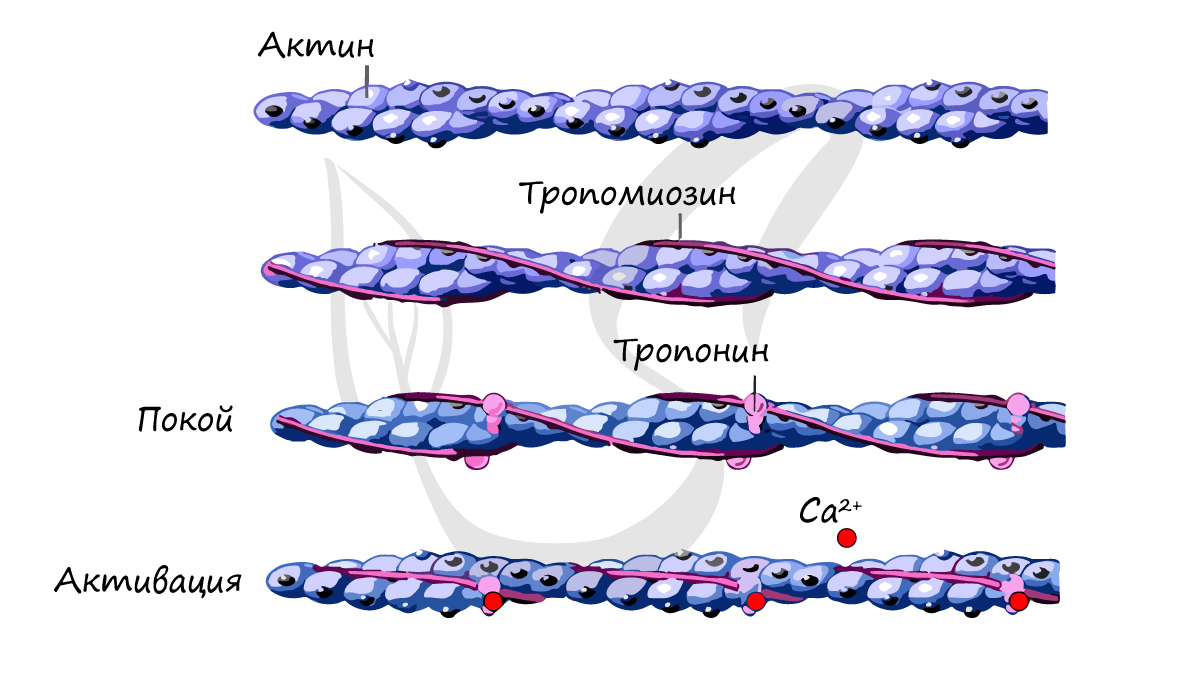 Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина. После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca). В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина. После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca). В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.  Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать. В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) . Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.
Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать. В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) . Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы. 
Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.  Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна. Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм. Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.
Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна. Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм. Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.  Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда. Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется. На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).
Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда. Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется. На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).  В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма. Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма. Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.  В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.
В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.  Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом. В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).
Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом. В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы). 
Происхождение мышц

Мышцы развиваются из среднего зародышевого листка — мезодермы. © Беллевич Юрий Сергеевич 2018-2021 Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию. Источник
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом