
В своих работах советский ученый Северцов А.Н. выделил понятия биологического прогресса и регресса.
Биологический прогресс подразумевает победу вида в борьбе за существование. Биологический прогресс характеризуется следующими признаками:
- Численность вида увеличивается
- Ареал расширяется
- Смертность особей уменьшается
- Рождаемость увеличивается
- Происходит процветание вида
Основными направлениями биологического прогресса являются:
-
Ароморфоз (греч. airomorphosis — поднимаю форму)
Ароморфоз представляет собой прогрессивное эволюционное преобразование, повышающее уровень организации организмов. В результате ароморфоза становится возможным освоение новых, ранее недоступных для жизни, территорий. К примеру, теплокровность птиц позволила им заселить места с холодным климатом.

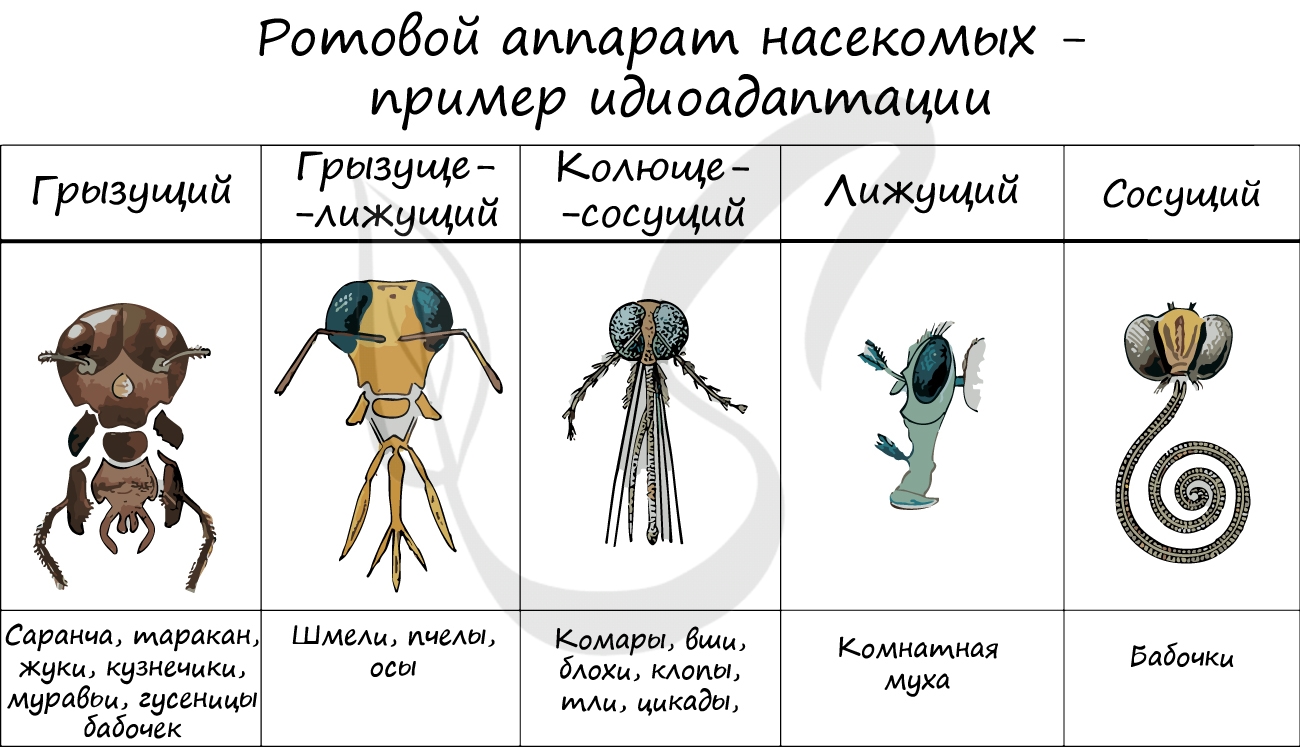
Идиоадаптация подразумевает незначительные, частные изменения в строении и функциях организма, которые помогают приспособиться к условиям среды обитания. Идиоадаптации существенно не повышают уровень организации.
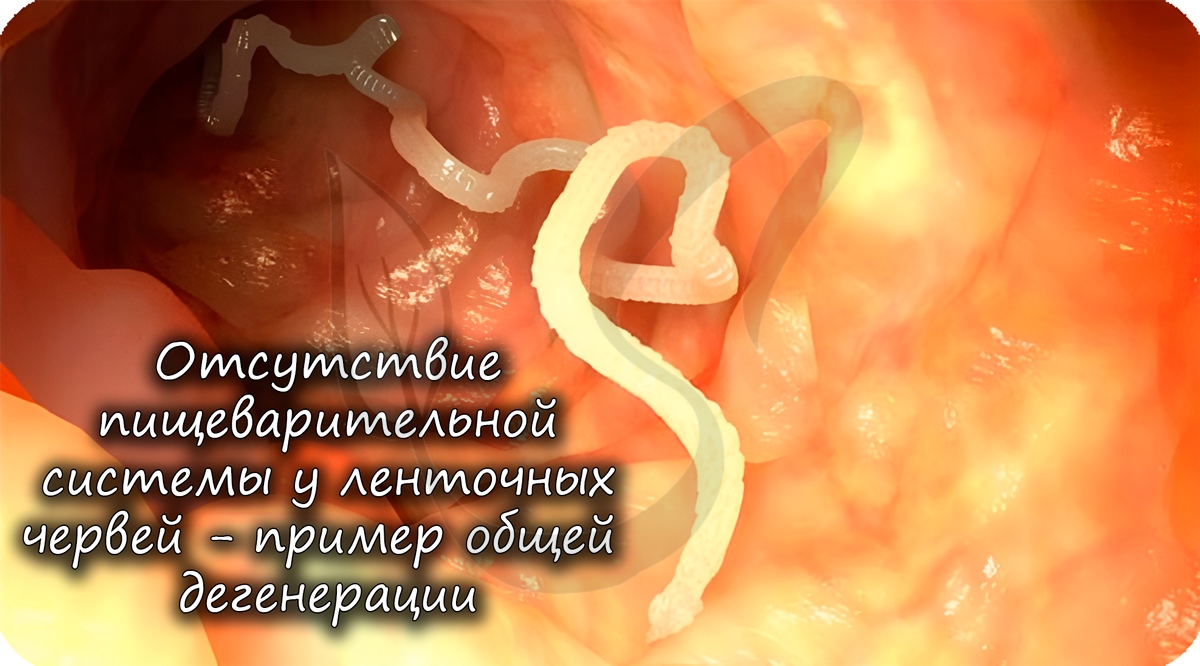
Общей дегенерацией называют упрощение организации, которое заключается в утрате отдельных органов и систем органов. У многих этот пункт вызывает внутреннее противоречие: как общая дегенерация может относиться к биологическому прогрессу?
На самом деле, если орган или система органов не нужна организму в его условиях обитания — то зачем она? Эта система может исчезнуть и освободить место для других, более полезных в данных условиях, органов.
У многих паразитов отсутствуют различные органы, к примеру, у ленточных червей нет пищеварительной системы. А зачем она им, когда пища в кишке, где они обитают, уже переварена и расщеплена организмом хозяина?
Биологический регресс характеризуется признаками, противоположными биологическому прогрессу:
- Численность вида уменьшается
- Ареал сужается
- Смертность особей возрастает
- Рождаемость уменьшается
- Происходит вымирание вида
Главная причина биологического регресса в том, что скорость эволюции вида отстает от скорости изменения внешней среды, эволюции других видов: это несоответствие снижает приспособленность организмов. Часто деятельность человека молниеносно меняет окружающую среду: далеко не все виды могут приспособиться к этому, происходит вымирание.

Сравнительно-анатомические доказательства эволюции
Изучение строения органов и их эволюционных изменений у различных групп организмов является основой выявления сравнительно-анатомических доказательств эволюции. Яркими примерами анатомических доказательств эволюции являются гомологичные и аналогичные органы.
-
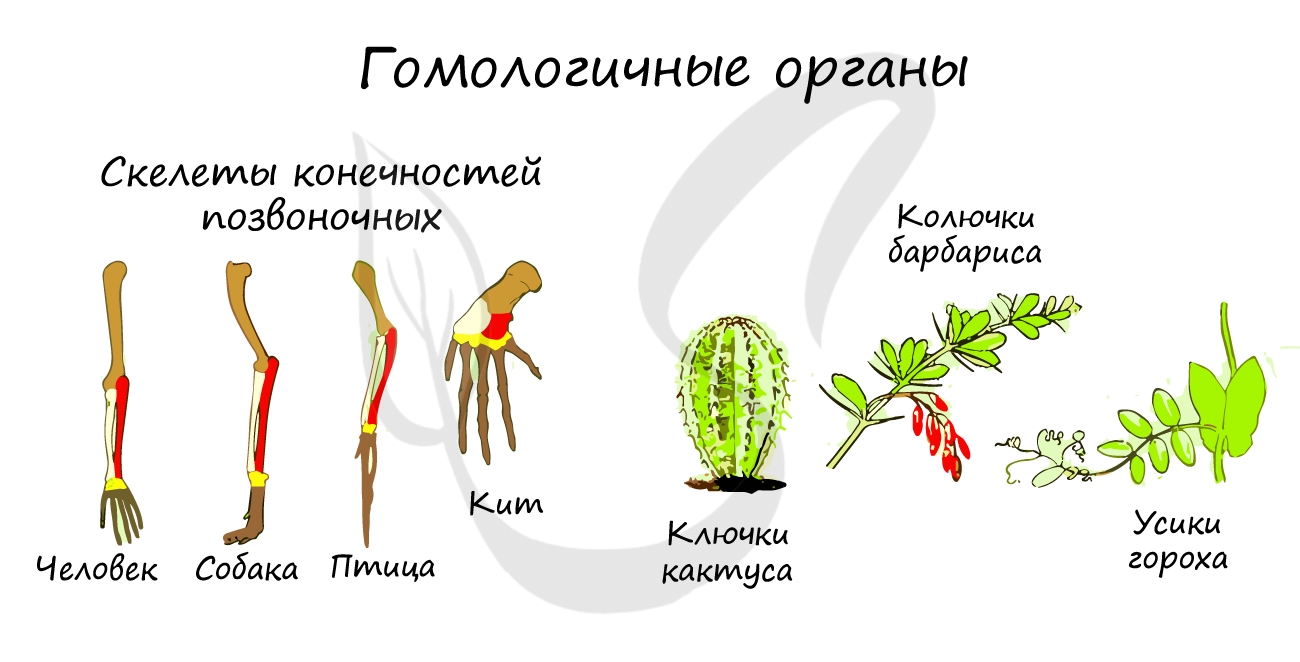
Гомологичные органы (гомология, от греч. homo(s) — равный, одинаковый)
Такие органы развиваются из одних и тех же зародышевых листков, имеют общий план строения, но выполняют разные функции. Это связано с тем, что животные освоили разные среды обитания, из-за чего происходит дивергенция (лат. divergo — отклоняюсь) — расхождение признаков у первоначально близких животных в ходе эволюции.
Гомологичны между собой скелеты конечностей различных классов позвоночных: рука — ласт — крыло птицы, колючки кактуса — усики гороха — листья растений.
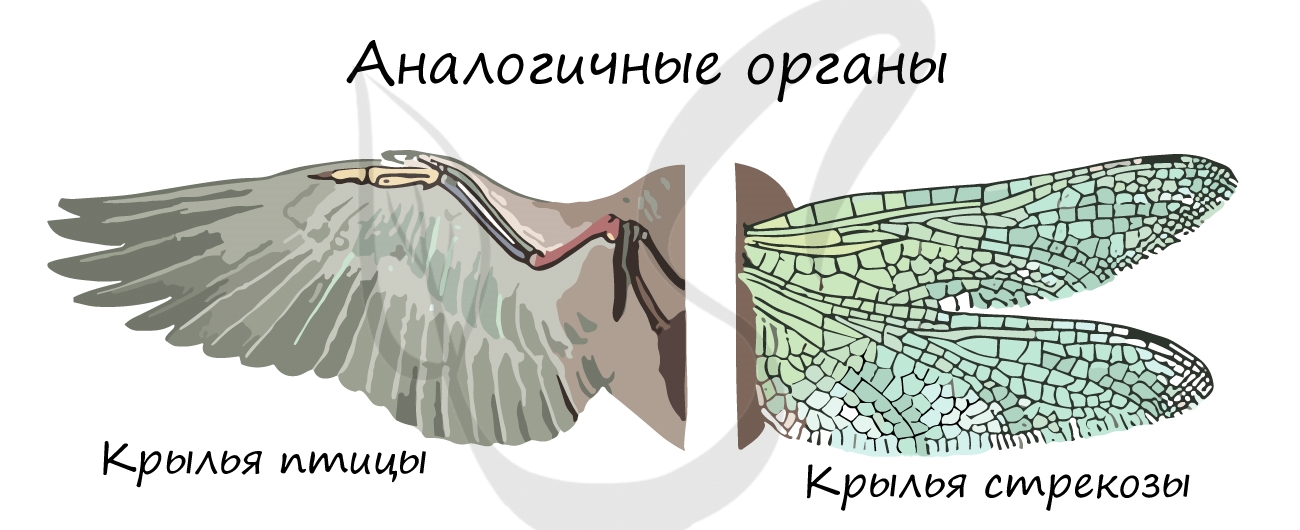
Аналогичные органы развиваются из разных зародышевых листков, имеют различное строение, но выполняют схожие функции. Такое сходство возникает в результате приспособления к одним и тем же условиям среды, из-за чего происходит конвергенция (лат. convergo — сближаю) — схождение признаков у неблизкородственных видов в ходе эволюции.
Аналогичными органами являются крыло птицы — крыло бабочки, глаз человека — глаз кальмара, усики винограда — усики гороха, жабры рака — жабры рыбы.
В строении нынешних животных можно найти признаки древних предковых форм, которые также свидетельствуют об эволюции. Сейчас мы обсудим рудименты и атавизмы.
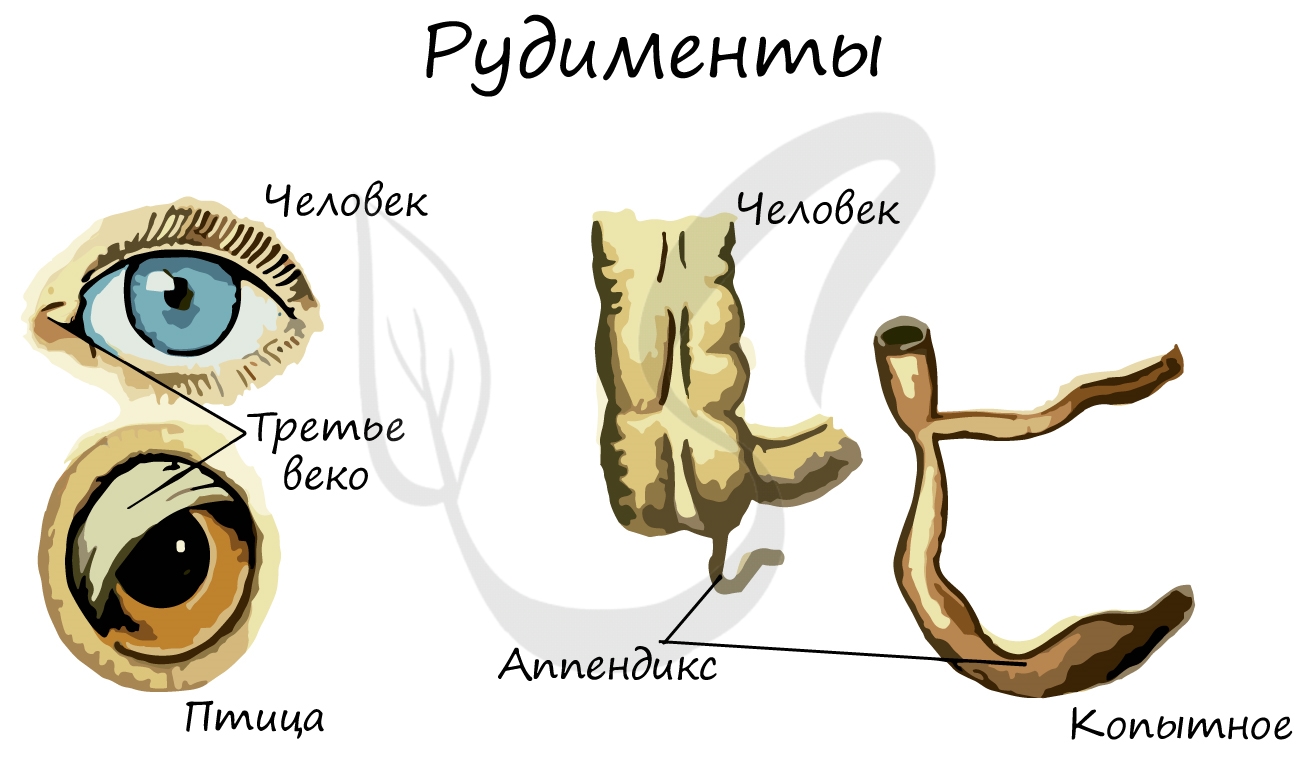
Рудименты (лат. rudimentum — зачаток) — органы, которые в ходе эволюции утратили свое функциональное значение. Они сохраняются в течение всей жизни и в норме обнаруживаются у человека и животных.
У человека к рудиментарным органам относятся: зубы мудрости, копчик, ушные мышцы, аппендикс (червеобразный отросток), третье веко (эпикантус).
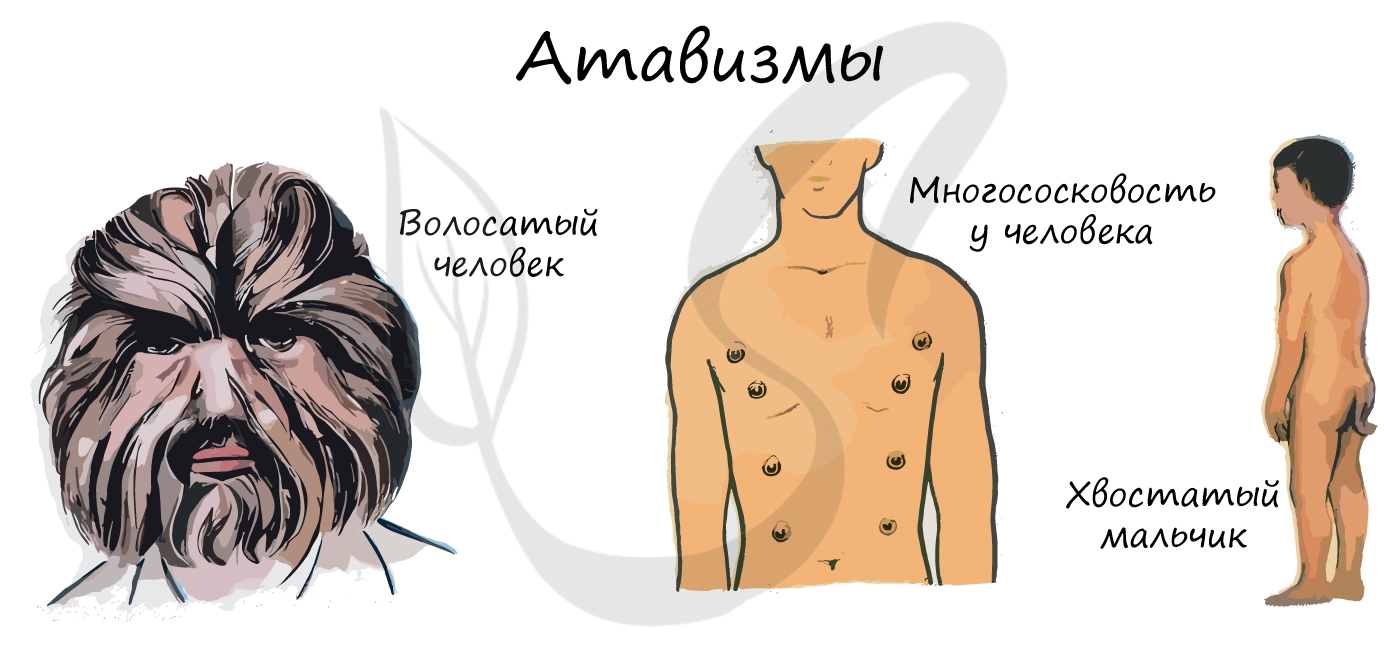
Атавизмы (лат. atavus — отдалённый предок) — случаи проявления у отдельных особей признаков дальних предков. Атавизмы сугубо индивидуальны и не являются нормой. Они также являются доказательством эволюции.
У человека атавизмами могут являться хвост, волосатое тело, добавочные молочные железы, незаращение межпредсердной перегородки.
Переходные формы
Переходные формы свидетельствуют о филогенетической преемственности, соединяя в своем строении черты высших и низших классов. Они — наглядное, живое доказательство эволюции.
Такими формами являются, к примеру, утконос и ехидна из класса млекопитающих. При многих признаках млекопитающих, они откладывают яйца, тем самым подтверждают родство млекопитающих с пресмыкающимися.

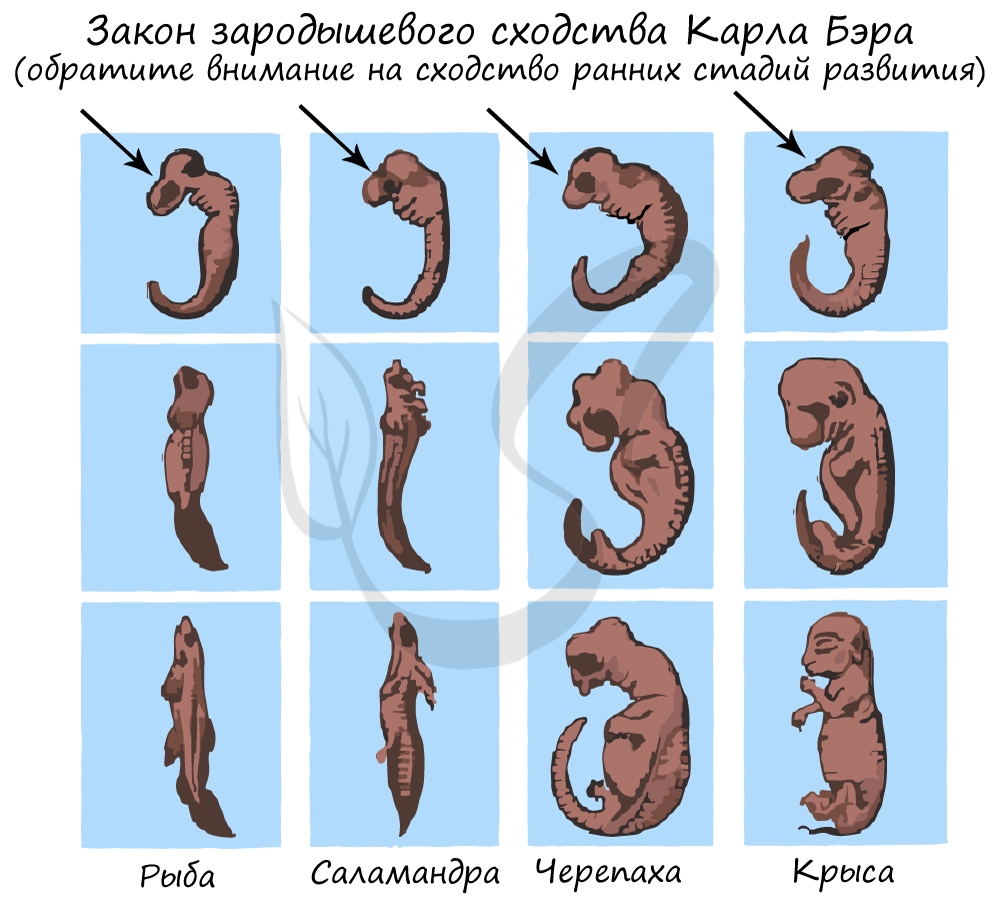
Эмбриологические доказательства
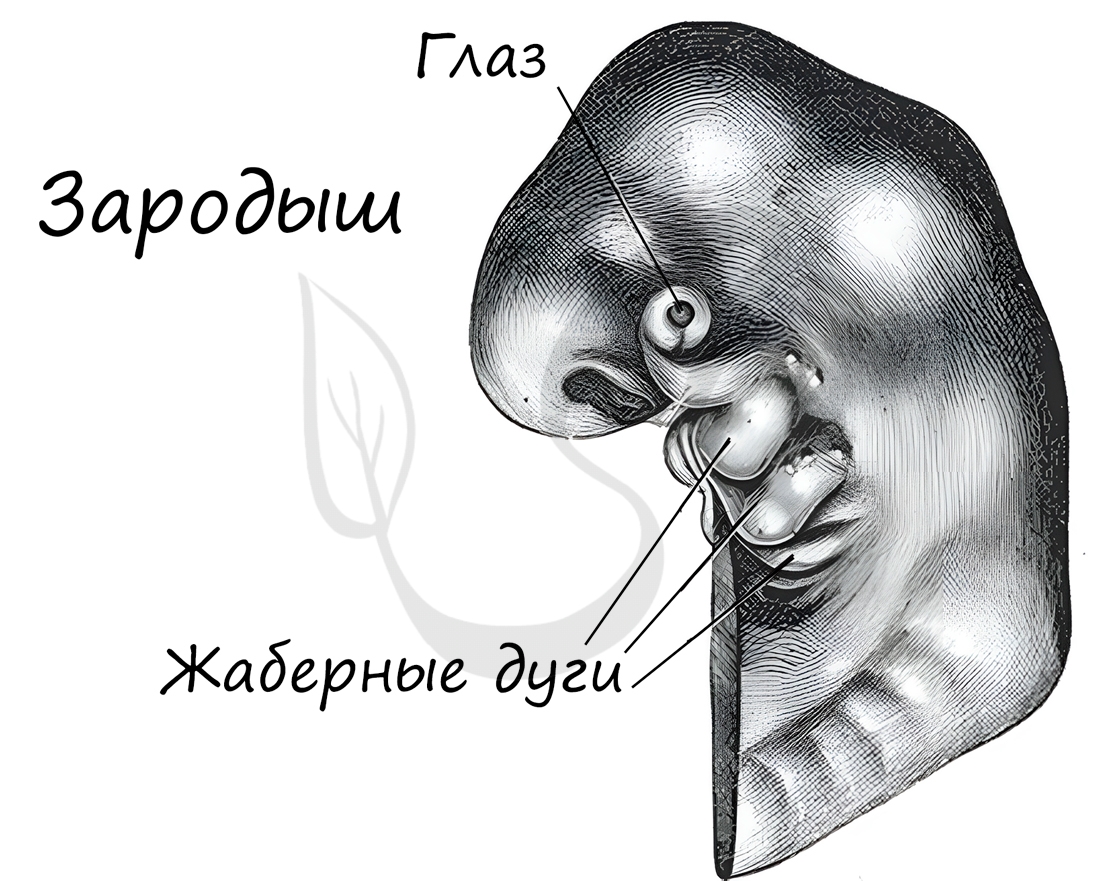
Эмбриология (греч. embryon — зародыш) — раздел биологии, изучающий строение эмбрионов. Только вдумайтесь: на этапе эмбриона, через который мы с вами успешно прошли, у нас можно было найти закладку жаберных дуг, которые существуют непродолжительное время, после чего исчезают.
А у рыб, например, жаберные дуги не исчезают — из них развиваются жабры.
Немецкие ученые Ф. Мюллер и Э. Геккель во второй половине XIX века сформулировали биогенетический закон, гласящий, что онтогенез (индивидуальное развитие) каждой особи есть краткое и быстрое повторение филогенеза (исторического развития вида).
Биогенетический закон Мюллера-Геккеля объясняет повторение этапов (на стадии зародыша), которые были свойственны нашим далеким предкам. Таким образом, мы проходим их этапы, но, не останавливаясь на них, двигаемся дальше к более совершенным этапам.
У головастиков лягушек развивается плавник, есть жабры — это наглядное повторение признаков, которые характерны для их предков — рыб.
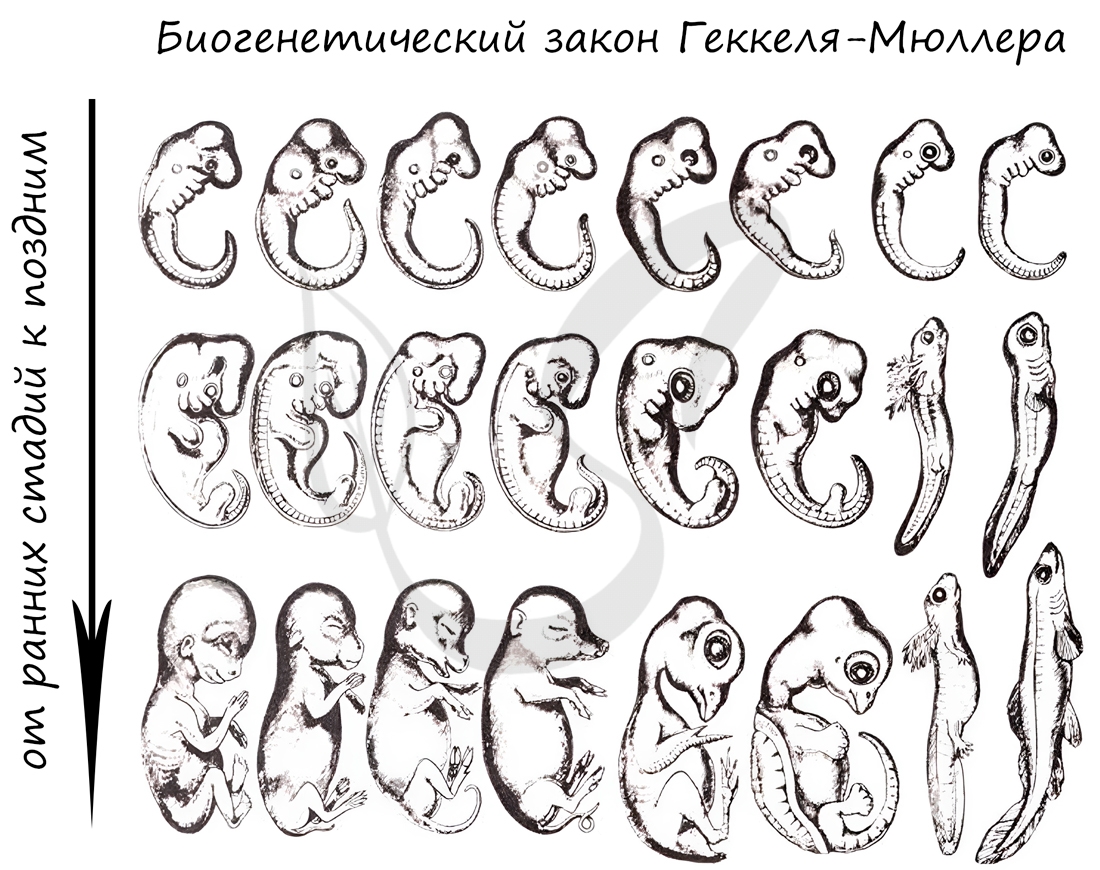
Карл Бэр сформулировал закон зародышевого сходства, который гласит, что на ранних стадиях развития зародыши позвоночных животных настолько похожи друг на друга, что практически неразличимы между собой. Это также указывает и подтверждает единство происхождения животного мира.
Читайте также: Законы функционирования возбудимых тканей физиология
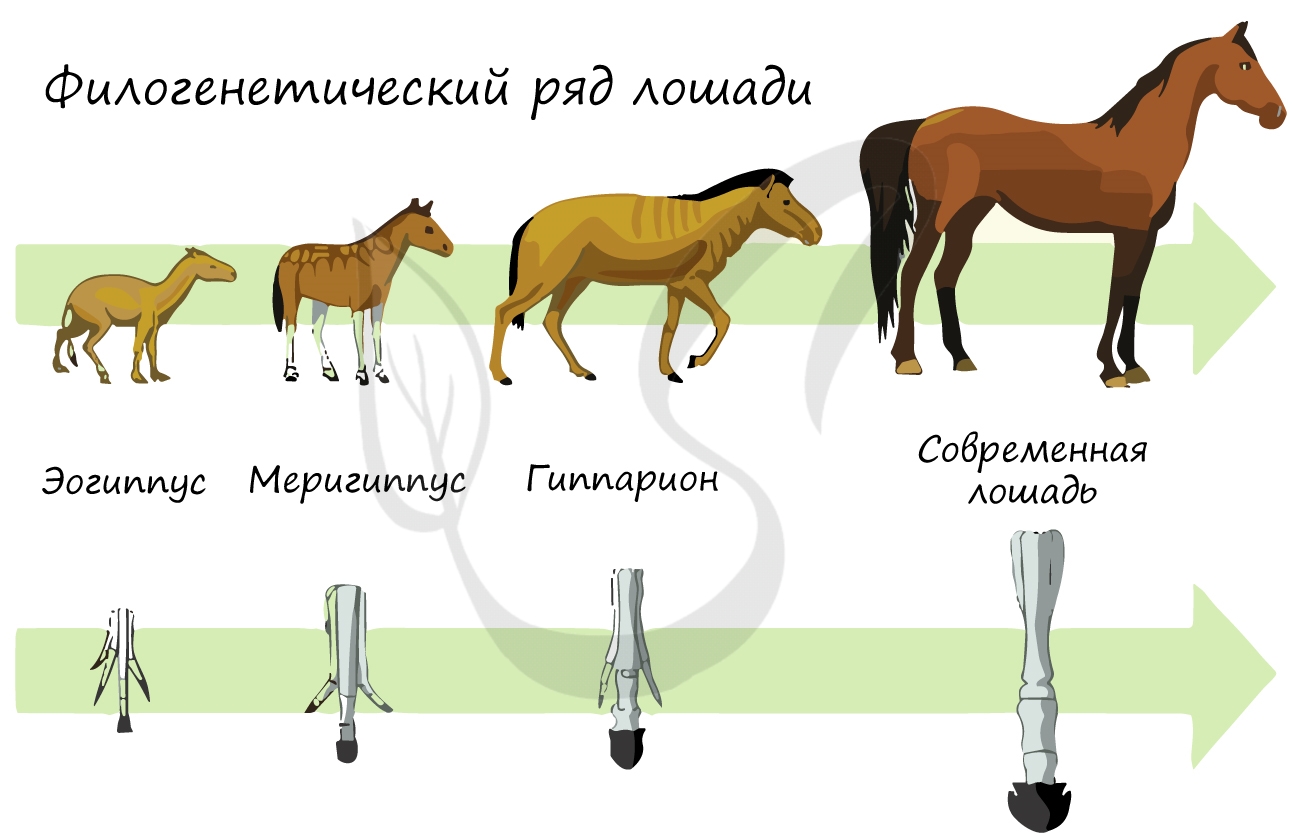
Палеонтологические доказательства эволюции
Палеонтология (греч. palaios – древний) изучает ископаемые останки вымерших животных, их сходства и различия с ныне живущими видами. Сопоставляя друг с другом ископаемые останки разных геологических эпох, можно увидеть как происходила эволюция различных видов животных и растений.
В результате таких исследований иногда удается открыть переходные формы, а иногда — целые филогенетические ряды, то есть совокупность последовательно сменяющих друг друга форм одного вида. Так, к примеру, был открыт филогенетический ряд лошади.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
ТЕОРИЯ ЭВОЛЮЦИИ ТКАНЕЙ
Ткани развиваются путем гистогенеза.
Гистогенез — единый комплекс координированных во времени и пространстве процессов пролиферации, дифференцировки, детерминации, интеграции и функциональной адаптации клеточных систем.
Развитие организма начинается с одноклеточной стадии — зиготы. В ходе дробления возникают бластомеры, но совокупность бластомеров – это еще не ткань. Бластомеры на начальных этапах дробления еще не детерминированы (они тотипотентны). Если отделить их один от другого, — каждый может дать начало полноценному самостоятельному организму – механизм возникновения монозиготных близнецов. Постепенно на следующих стадиях происходит ограничение потенций. В основе его лежат процессы, связанные с блокированием отдельных компонентов генома клеток и детерминацией.
Детерминация – это процесс определения дальнейшего пути развития материала эмбриональных зачатков с образование специфических тканей (на основе блокирования отдельных генов).
Понятие «коммитирование» тесно связано с клеточным делением (т.н. коммитирующий митоз).
Коммитирование – это ограничение возможных путей развития вследствие детерминации. Коммитирование совершается ступенчато. Сначала соответствующие преобразования генома касаются крупных его участков. Затем все более детализируются, поэтому вначале детерминируются наиболее общие свойства клеток, а затем и более частные.
Как известно, на этапе гаструляции возникают эмбриональные зачатки. Клетки, которые входят в их состав, еще не окончательно детерминированы, так что из одного зачатка возникают клеточные совокупности, обладающие разными свойствами. Следовательно, один эмбриональный зачаток может служить источником развития нескольких тканей.
Последовательная ступенчатая детерминация и коммитирование потенций однородных клеточных группировок — дивергентный процесс. В общем виде эволюционная концепция дивергентного развития тканей в филогенезе и в онтогенезе была сформулирована Н.Г.Хлопиным. Современные генетические концепции подтверждают правоту его представлений. Именно Н.Г.Хлопин ввел понятие о генетических тканевых типах. Концепция Хлопина хорошо отвечает на вопрос, как и какими путями происходило развитие и становление тканей, но не останавливается на причинах, определяющих пути развития.
Причинные аспекты развития тканей раскрывает теория параллелизмовА.А.Заварзина. Он обратил внимание на сходство строения тканей, которые выполняют одинаковые функции у животных, принадлежащих даже к весьма удаленным друг от друга эволюционным группировкам. Вместе с тем известно, что, когда эволюционные ветви только расходились, у общих предков таких специализированных тканей еще не было. Следовательно, в ходе эволюции в разных ветвях филогенетического древа самостоятельно, как бы параллельно, возникали одинаково организованные ткани, выполняющие сходную функцию. Причиной этого является естественный отбор: если возникали какие-то организмы, у которых соответствие строения и функции клеток, тканей, органов нарушалось, они были и менее жизнеспособны. Теория Заварзина отвечает на вопрос, почему развитие тканей шло тем, а не иным путем, раскрывает казуальные аспекты эволюции тканей.
Концепции А.А.Заварзина и Н.Г.Хлопина, разработанные независимо одна от другой, дополняют друг друга и были объединены А.А.Брауном и В.П.Михайловым: сходные тканевые структуры возникали параллельно в ходе дивергентного развития.
Развитие тканей в эмбриогенезе происходит в результате дифференцировки клеток. Под дифференцировкой понимают изменения в структуре клеток в результате их функциональной специализации, обусловленные активностью их генетического аппарата. Различают четыре основных периода дифференцировки клеток зародыша — оотипическую, бластомерную, зачатковую и тканевую дифференцировку. Проходя через эти периоды, клетки зародыша образуют ткани (гистогенез).
Эволюция клеток и тканей
Происхождение, специализация и объединение клеток в ткани есть категории исторические, ибо они возникли в ходе филогенеза. Однако объяснение этих категорий чрезвычайно затруднено, поскольку между клетками прокариотических и эукариотических организмов существуют важные различия. Тем не менее известно несколько гипотез.
На основании изучения ископаемых остатков бактерий и циано-бактерий предполагают, что предковой клеточной формой была примитивная прокариотическая клетка, возникшая около 3,5 х 10 9 лет назад. Клетки этого типа для обеспечения своего существования и размножения в начале использовали органические молекулы небиологического происхождения. Первым актом в формировании примитивных клеток было образование мембраны, окружавшей вещество клетки.
В последующем у примитивных прокариотических клеток стали развиваться механизмы синтеза и энергетического обеспечения. Предполагают, что первые прокариотические клетки обладали наиболее простыми каталитическими системами, в результате чего обеспечение их энергией основывалось на брожении. В последующем отдельные виды прокариотических клеток переключились с брожения на дыхание, что способствовало более эффективному получению энергии. Таким образом, эволюционные изменения прокариотических клеток шли по линии развития у них различных метаболических путей. Их геном развивался в направлении формирования «голых» молекул ДНК.
Читайте также: Американка шов для легкой прозрачной ткани пошагово
Эволюционные изменения эукариотических клеток шли в направлении увеличивающегося разнообразия в форме, размерах, структуре и функциях с одновременной компартментализацией биохимических систем и сохранением общего для всех Клеток аэробного метаболизма. Считают, что эукариотические клетки возникли менее 1 млрд лет назад из прокариотических клеток, причем для объяснения их происхождения выдвинуто три гипотезы.
В соответствии с одной их этих гипотез (гипотезой клеточного симбиоза), которая является наиболее распространенной, предполагают, что эукариотическая клетка является симбиотической структурой, состоящей из нескольких клеток разных типов, объединенных общей клеточной мембраной. В частности, предполагают, что пластиды клеток современных зеленых растений происходят от бактерий, бывших предками современных цианобактерий и способных к аэробному фотосинтезу, а митохондрии эукариотических клеток ведут начало от аэробных бактерий, которые вступали в симбиоз с примитивными анаэробными клетками, способными к фотосинтезу, что привело к образованию клеток, способных к существованию в атмосфере кислорода и использованию кислорода путем дыхания. Относительно ядра предполагают, что оно является рудиментом также какого-то древнего внутриклеточного симбионта, утратившего после включения в исходную клетку свою цитоплазму. В пользу этой гипотезы свидетельствуют данные о симбиотических взаимоотношениях некоторых современных организмов. Например, одноклеточная зеленая водоросль хлорелла (Chlorella) обитает в цитоплазме зеленой парамеции (Paramecium bussaria). Из-за способности к фотосинтезу она снабжает парамеции питательными веществами. Пластиды и митохондрии содержат собственную систему генетической информации о синтезе белков в виде ДНК, мРНК, рРНК, тРНК и соответствующих ферментов. Для хлоропластов, митохондрии и клеток-прокариот характерно сходство способов репродукции (все они одинаково репродуцируются путем простого деления надвое). Наконец, мутации митохон-дриальных генов назависимы от мутаций ядерных генов.
В соответствии с другой гипотезой считают, что эукариотичес-кая клетка произошла от прокариотической клетки, содержавшей несколько геномов, прикрепленных к клеточной мембране. В результате инвагинаций клеточной мембраны образовывались мезо-сомы, способные первоночально к фотосинтезу. Однако в дальнейшем произошла специализация этих органелл, в результате чего одна из них, утратив дыхательную и фотосинтетическую функцию, развилась в ядро, другие, наоборот, развив эти функции, стали митохондриями у животных и пластидами у растений. В пользу этой гипотезы свидетельствуют данные о двойном строении мембран ядра, митохондрии и пластид.
В соответствии с третьей гипотезой, основанной на мысли о том, что все живые формы произошли от предковых анаэробных ферментативных гетеротрофов, эукариоты представляют собой сублинию бесстеночных (анаэробных) прокариотов, которые развили способность к эндоцитозу. Посредством «заглатывания» других прокариотов, которые дали им дополнительные метаболические способности и которые, в конце концов, дегенерировали в органеллы, примитивная клетка (уркариот) стала эукариотической клеткой. Таким образом, прокариоты древнее, проще и примитивнее клеток-эукариот.
В соответствии с четвертой гипотезой предполагают, что эука-риотические клетки возникли из прокариотической клетки, содержавшей много геномов, которые распадались на части, давшие начало структурам с разными функциями. В последующем шло кло-нирование структур со сходными функциями, после чего они покрывались двойными мембранами, что привело к образованию ядра, митохондрии, а позднее и мембранной сети. В пользу этой гипотезы свидетельствуют данные о сходстве генетического кода, содержащегося в ядерной и митохондриальной ДНК, а также о сходстве в регуляции дыхательной функции ядром и митохондриями.
Как отмечено выше, симбиотическая гипотеза происхождения эукариотических клеток сейчас наиболее популярна. Однако, разделяя эту гипотезу, нельзя не отметить, что митохондрии и хло-ропласты вопреки их сходству с современными бактериями-аэробами и цианобактериями (соответственно) все же существенно отличаются от них. В частности, в митохондриях и хлоропластах намного меньше ДНК. Следовательно, здесь митохондрии и хлоропласты в ходе эволюции претерпели значительные изменения в направлении своих размеров.
Геном эукариотических клеток впоследствии развивался в направлении объединения молекул ДНК с белками и формирования хроматина и хромосом разной формы и в разном количестве. Специализация хроматина проявилась в формировании эухроматина и гетерохроматина, в формировании аутосом и половых хромосом. Что касается количества хромосом, то объяснить их эволюционную тенденцию пока трудно, поскольку многие примитивные организмы содержат в своих клетках большее число хромосом, чем организмы, занимающие высшие эволюционные ступени. Однако несомненно, что количественные и структурные изменения карио-типов в течение эволюции играли важную роль в видообразовании. Параллельно с этим происходило усложнение структуры и функции клеточных компонентов, развитие регуляторных механизмов.
Несомненно эволюционное значение митоза. Считают, что точность разделения и распределения хромосом в результате митоза является условием, обеспечивающим многоклеточность. Однако происхождение самого митоза не имеет достаточных объяснений. Предполагают лишь, что он развился из примитивного митоза, представляющего собой механизм, при котором расхождение реп-лицировавшихся хромосом происходило после вытягивания и разрыва веретена без разрушения ядерной мембраны (см. выше).
Объяснения эволюции тканей связаны со сложностями, которые обусловлены одинаковым строением тканей, принадлежащих живым организмам, находящимся на разных ступенях эволюционной лестницы. Например, мышечные волокна членистоногих, некоторых моллюсков и позвоночных имеют одинаковое строение. Между тем эти организмы филогенетически разделены очень большими «расстояниями». Аналогичная ситуация имеет место и при сравнении тканей растений из разных таксономических групп.
Читайте также: Шторы из ткани канвас коричневый
Начала тканеобразования в эволюционном плане уже прослеживаются у самых простых организмов. Например, у вольвокса отмечается формирование колоний, состоящих иногда более чем из 50 000 клеток, причем часть клеток уже специализирована. В частности, клетки, располагающиеся по краям колониальной формы, ответственны за образование новых колоний. У цианобактерий при нерасхождении разделившихся клеток образуются клеточные нити, в которых часть клеток специализирована на фиксации азота, чем обеспечиваются потребности в азоте и других клеток.
Идя вверх по эволюционной лестнице, можно видеть, что у губок уже отмечается около пяти специализированных типов клеток, специализация которых связана с выполнением разных функций в процессе фильтрации воды и поглощения отфильтрованных пищевых частиц.
У кишечнополостных тело состоит из двух слоев — эктодермы и энтодермы, представляющих собой наружный и внутренний эпителиальные слои. Наружные эпителиальные клетки являются стрекательными клетками, содержащими ядовитую жидкость, тогда как внутренние эпителиальные клетки секретируют пищеварительные ферменты и обеспечивают пищеварение. Поэтому предполагают, что первыми сформировались слои эпителиальных клеток и их роль в эволюции многоклеточных аналогизируется с ролью клеточных стенок и мембран одноклеточных организмов.
Значительный вклад в понимание эволюции тканей принадлежит А. А. Заварзину (1886-1945), который считал, что одни и те же факторы эволюции обеспечили не только разнообразие организмов, но и однообразие строения их тканей. Сходство в строении тканей у филогенетически далеко отстоящих животных А. А. За-варзин называл законом параллельных рядов тканевой эволюции. Работы А. А. Заварзина и его учеников заложили основы эволюционной гистологии.
Вопросы для обсуждения
1. Насколько велико значение методов исследования в изучении клеток? Какие из этих методов вы знаете?
2. Сформулируйте основные положения клеточной теории. Как Вы считаете, какова роль этой теории в биологии?
3. Почему клетку определяют в качестве элементарной единицы жизни и в чем заключаются доказательства того, что клетка действительно является элементарной единицей жизни? Что представляют собой межклеточные структуры?
4. Назовите два процесса, которые являются общими для всех живых систем.
б. Назовите принципиальные различия между клетками-прокариота-ми и клетками-эукариотами. Является ли одноклеточность признаком прокариот?
6. Назовите и охарактеризуйте компоненты мембранной системы клеток животных. Есть ли мембранная система в клетках растений?
7. Охарактеризуйте цитоплазматический матрикс и клеточные орга-неллы. Что собой представляет цитозоль? Есть ли у клеток скелет? Как организован цитоскелет и каковы его компоненты?
8. Каковы структура и роль клеточного ядра? Есть ли различия между ядрами клеток животных и клеток растений?
9. Каковы структура и функции митохондрий? Все ли клетки обладают митохондриями?
10. Сформулируйте определения клеточного цикла и митоза. С какой скоростью протекают митозы в клетках разных тканей?
11. Что собой представляют лизосомы и какова их роль? Что произойдет с клетками, если лизосомы подвергнутся разрушению?
12. Каково значение ферментов в жизни клеток? Все ли белки являются ферментами и в чем заключается их действие?
13. Каковы фазы митоза и сущность процессов, происходящих в эти фазы?
14. В какой фазе происходит разделение центромеры и освобождение сестринских хроматид?
15. Определите, какая весовая часть ядра приходится на хроматин клетки (примерно), допуская, что диаметр ядра составляет б мкм, а плотность 1,1 г/см» 3 ?
16. Считая, что хромосомы человека состоят на 15% из ДНК, определите массу всех хромосом его диплоидных клеток.
17. Что вы можете сказать о происхождении митоза?
18. Что вы знаете об элементарном составе клеток?
19. Что понимают под биологическими молекулами?
20. Какой представляется вструктура белков и что вы знаете о функциях белков?
21. Как вы понимаете происхождение клеток-прокариот?
22. Как вы понимаете происхождение клеток-эукариот?
23. Каково ваше мнение относительно развития генома эукариотичес-ких клеток?
24. Каковы причины гибели клеток? Существует ли генетический механизм, контролирующий гибель клеток?
25. Что вы знаете об эпителиальных тканях и их функциях?
26. Назовите основные группы мышечной ткани и что составляет основу их классификации?
27. Каковы основные клеточные элементы собственно соединительных тканей?
28. Что такое нервная ткань и из каких компонентов она состоит?
29. Как вы представляете строение нервного волокна?
30. Почему кровь и лимфу считают тканями?
31. Какова функциональная роль лимфоцитов?
32. Как вы понимаете происхождение клеток-прокариотов и клеток-эукариотов?
33. Применима ли эволюционная теория к учению о тканях?
Альберте В., Брей Д., Льюис Дж„ Рэфф М., Роберте К., Уотсон Дж. Молекулярная биология клетки. М.: Мир. 1994. Т. 1. 615 стр.; 1994. Т. 2. 540 стр.
Вермель Е. М. История учения о клетке. М.: Наука. 1970. 259 стр.
Kaufman Р. В., Wu W. Handbook of Molecular and Cellular Methods in Biology and Medicine. CRC Press L. 1995. 496 pp. Lackie J. M„ Dow 3. A. The Dictionary of Cell Biology. Academic Press. 1995. 380 pp.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности