
2.2.1. Аммиак (NН3) – продукт обмена большинства соединений, содержащих амино- и амидогруппы. Главным путём образования аммиака служит окислительное дезаминирование.
Аммиак – очень токсичное вещество, особенно для нервной системы. При физиологических значениях рН молекула NН3 легко превращается в ион аммония NН4 + , который не способен проникать через биологические мембраны и задерживается в клетке. Накопление NН4 + вызывает торможение заключительных этапов цикла трикарбоновых кислот и снижение продукции АТФ. Поэтому в организме существует ряд механизмов связывания (обезвреживания) аммиака (см. рисунок 2.3).
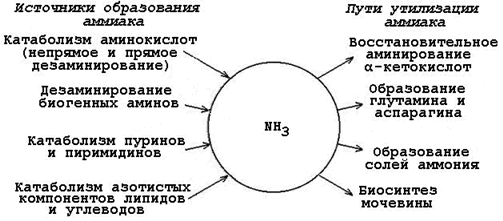
Рисунок 2.3. Образование аммиака в организме и его обезвреживание.
2.2.2. Образование транспортных форм аммиака в тканях. Аммиак, образующийся в тканях, сначала превращается в нетоксичное соединение и в таком виде переносится кровью к печени или почкам. Такими транспортными формами являются аминокислоты глутамин, аспарагин и аланин.
Образование глутамина и аспарагина из глутамата и аспартата соответственно происходит во многих тканях, включая головной мозг:


Глутамин — нейтральное нетоксичное соединение, способное легко проходить через клеточные мембраны. В виде этой аминокислоты аммиак транспортируется в крови. В крови здоровых людей содержание глутамина существенно превышает содержание других аминокислот. Глутамин, помимо участия в синтезе белка, служит источником азота в биосинтезе гистидина, глюкозамина, пуриновых и пиримидиновых нуклеотидов. С кровью глутамин поступает в печень и почки. Здесь он под действием фермента глутаминазы превращается в глутамат и аммиак. При участии аспарагиназы также происходит образование аммиака из аспарагина.
Аланин является транспортной формой аммиака, которая образуется преимущественно в мышцах. При интенсивной физической нагрузке источниками аммиака служат реакции дезаминирования аминокислот и аденозинмонофосфата (АМФ). Сначала аммиак превращается в аминогруппу глутамата в реакции восстановительного аминирования (см. параграф 2.1.2), катализируемой глутаматдегидрогеназой:

Образовавшийся глутамат переносит затем свою α-аминогруппу на пируват, всегда имеющийся в достаточном количестве, поскольку это продукт протекающего в мышцах гликолиза. Реакция катализируется аланинаминотрансферазой.

Глутамат + Пируват α-Кетоглутарат + Аланин
Аланин (нейтральная аминокислота, не несущая суммарного заряда при значениях рН, близких к 7) выходит из клеток и доставляется кровью к печени. Здесь он под действием аланинаминотрансферазы передаёт свою аминогруппу α-кетоглутарату, в результате чего образуется глутамат.

α-Кетоглутарат + Аланин Глутамат + Пируват
Далее в реакции, катализируемой глутаматдегидрогеназой, глутамат дезаминируется с образованием α-кетоглутарата и аммиака, который в печени превращается в мочевину.
2.2.3. Экскреция аммиака с мочой. Клетки почек поглощают из циркулирующей крови глутамин. Фермент глутаминаза в почках катализирует гидролиз глутамина с образованием глутамата и аммиака.

Образующийся глутамат может в дальнейшем подвергаться дезаминированию при участии глутаматдегидрогеназы. Таким образом, из одной молекулы глутамина всего может образоваться две молекулы аммиака.
Аспарагин аналогичным образом подвергается гидролизу при участии аспарагиназы, присутствующей в клетках канальцев почек.
Аммиак секретируется в просвет почечных канальцев и выводится в виде солей минеральных и органических кислот, например:

NН3 + НСl NН4Сl хлорид аммония
Образование солей аммония в почечных канальцах является важным механизмом регуляции кислотно-основного состояния организма. Оно резко возрастает при метаболическом ацидозе — накоплении в организме кислот и снижается при потере кислот организмом (алкалозе).
2.2.4. Биосинтез мочевины – основной путь обезвреживания аммиака. Мочевина синтезируется в орнитиновом цикле, протекающем в клетках печени. Эту последовательность реакций открыли Х.Кребс и К.Хензелейт в 1932 г. Согласно современным представлениям, цикл мочевины включает последовательность пяти реакций.
Читайте также: Зернистая масса продукт распада тканей 6 букв
Две начальные реакции биосинтеза мочевины происходят в митохондриях клеток печени.


Последующие реакции протекают в цитоплазме клеток печени.



Общая схема орнитинового цикла представлена на рисунке 2.4:

Рисунок 2.4. Схема орнитинового цикла и его связь с превращениями фумаровой и аспарагиновой кислот. Цифрами обозначены ферменты, катализирующие реакции орнитинового цикла: 1 – карбамоилфосфатсинтетаза; 2 – орнитин-карбамоилтрансфераза; 3 – аргининосукцинатсинтетаза; 4 – аргининосукцинатлиаза; 5 – аргиназа.
2.2.5. Орнитиновый цикл находится в тесной взаимосвязи с циклом трикарбоновых кислот:
пусковые реакции цикла мочевины, как и реакции ЦТК, протекают в митохондриальном матриксе;
поступление СО2 и АТФ, необходимых для образования мочевины, обеспечивается работой ЦТК;
в цикле мочевины образуется фумарат, который является одним из субстратов ЦТК. Фумарат гидратируется в малат, который в свою очередь окисляется в оксалоацетат. Оксалоацетат может подвергаться трансаминированию в аспартат; эта аминокислота участвует в образовании аргининосукцината.
2.2.6. Регуляция активности ферментов цикла осуществляется главным образом на уровне карбамоилфосфатсинтетазы, которая малоактивна в отсутствие своего аллостерического активатора — N-ацетил-глутамата. Концентрация последнего зависит от концентрации его предшественников (ацетил-КоА и глутамата), а также аргинина, который является аллостерическим активатором N-ацетилглутаматсинтазы:

Ацетил-КоА + Глутамат N-ацетилглутамат + КоА-SH
Концентрация ферментов орнитинового цикла зависит от содержания белка в пищевом рационе. При переходе на диету, богатую белком, в печени повышается синтез ферментов орнитинового цикла. При возвращении к сбалансированному рациону концентрация ферментов снижается. В условиях голодания, когда усиливается распад тканевых белков и использование аминокислот как энергетических субстратов, возрастает продукция аммиака, концентрация ферментов орнитинового цикла увеличивается.
2.2.7. Нарушения орнитинового цикла. Известны метаболические нарушения, обусловленные частичным блокированием каждого из 5 ферментов, катализирующих в печени реакции синтеза мочевины, а также N-ацетилглутаматсинтазы. Эти генетические дефекты, очевидно, являются частичными. Полное блокирование какой-либо из стадий цикла мочевины в печени, по-видимому, несовместимо с жизнью, потому что другого эффективного пути удаления аммиака не существует.
Общим признаком всех нарушений синтеза мочевины является повышенное содержание NH4 + в крови (гипераммониемия). Наиболее тяжёлые клинические проявления наблюдаются при дефекте фермента карбамоилфосфатсинтетазы. Клиническими симптомами, общими для всех нарушений цикла мочевины, являются рвота, нарушение координации движений, раздражительность, сонливость и умственная отсталость. Если заболевание не диагностируется, то быстро наступает гибель. У детей старшего возраста проявлениями заболевания служат повышенная возбудимость, увеличение размеров печени и отвращение к пище с высоким содержанием белка.
Лабораторная диагностика заболеваний включает определение содержания аммиака и метаболитов орнитинового цикла в крови, моче и спинномозговой жидкости; в сложных случаях прибегают к биопсии печени.
Значительное улучшение наблюдается при ограничении белка в диете, при этом могут быть предотвращены многие нарушения мозговой деятельности. Малобелковая диета приводит к снижению содержания аммиака в крови и к улучшению клинической картины при мягких формах этих наследственных нарушений. Пищу следует принимать часто, небольшими порциями, для того чтобы избежать резкого повышения уровня аммиака в крови.
2.2.8. Клинико-диагностическое значение определения мочевины в крови и моче. В крови здорового человека содержание мочевины составляет 3,33 – 8,32 ммоль/л. За сутки с мочой выводится 20 – 35 г мочевины.
Читайте также: Мембранный потенциал в различных возбудимых тканях имеет величину в диапазоне
Изменения содержания мочевины в крови при заболеваниях зависят от соотношения процессов её образования в печени и выведения почками. Повышение содержания мочевины в крови (гиперазотемия) отмечается при почечной недостаточности, снижение – при недостаточности печени, при диете с низким содержанием белков.
Повышение экскреции мочевины с мочой наблюдается при употреблении пищи с высоким содержанием белков, при заболеваниях, сопровождающихся усилением катаболизма белков в тканях, при приёме некоторых лекарств (например, салицилатов). Снижение экскреции мочевины с мочой характерно для заболеваний и токсических поражений печени, заболеваний почек, сопровождающихся нарушением их фильтрационной способности.
Обезвреживание аммиака в организме
В организме человека подвергается распаду около 70 г аминокислот в сутки, при этом в результате реакций дезаминирования и окисления биогенных аминов освобождается большое количество аммиака, являющегося высокотоксичным соединением. Поэтому концентрация аммиака в организме должна сохраняться на низком уровне. Действительно, уровень аммиака в крови в норме не превышает 60 мкмоль/л (это почти в 100 раз меньше концентрации глюкозы в крови). В опытах на кроликах показано, что концентрация аммиака 3 ммоль/л является летальной. Таким образом, аммиак должен подвергаться связыванию в тканях с образованием нетоксичных соединений, легко выделяющихся с мочой.
Один из путей связывания и обезвреживания аммиака в организме, в частности в мозге, сетчатке, почках, печени и мышцах,– это биосинтез глутамина (и, возможно, аспарагина). Глутамин и аспарагин выделяются с мочой в небольшом количестве. Было высказано предположение, что они выполняют скорее транспортную функцию переноса аммиака в нетоксичной форме. Ниже приводится химическая реакция синтеза глутамина, катализируемого глутаминсинтетазой.
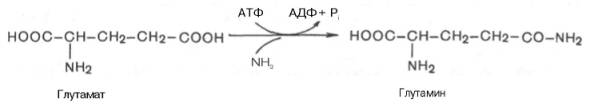
Механизм этой синтетазной реакции, подробно изученный А. Майсте-ром, включает ряд стадий. Синтез глутамина в присутствии глутамин-синтетазы может быть представлен в следующем виде:
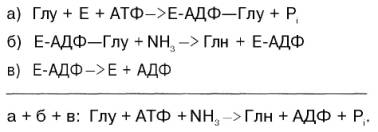
Биосинтез аспарагина протекает несколько отлично и зависит от природы ферментов и донора аммиака. Так, у микроорганизмов и в животных тканях открыта специфическая аммиакзависимая аспарагинсинтетаза, которая катализирует синтез аспарагина в две стадии:
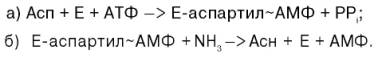
В животных тканях содержится, кроме того, глутаминзависимая аспа-рагинсинтетаза, которая для синтеза во второй стадии использует амидную группу глутамина:
Суммарная ферментативная реакция синтеза аспарагина может быть представлена в следующем виде:
Асп + АТФ + NН3 (или Глн) –> Асн + АМФ + РРi + (Глу).
Видно, что энергетически синтез аспарагина обходится организму дороже, поскольку образовавшийся РРi далее распадается на ортофосфат.
Часть аммиака легко связывается с α-кетоглутаровой кислотой благодаря обратимости глутаматдегидрогеназной реакции. Если учесть связывание одной молекулы аммиака при синтезе глутамина, то нетрудно видеть, что в организме имеется хорошо функционирующая система, связывающая две молекулы аммиака:
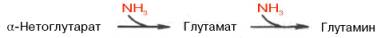
Глутамин, кроме того, используется почками в качестве резервного источника аммиака (образуется из глутамина под действием глутаминазы), необходимого для нейтрализации кислых продуктов обмена при ацидозе и защищающего тем самым организм от потери с мочой используемых для этих целей ионов Na + .
Роль глутамина и аспарагина в обезвреживании аммиака.
1. Основная реакция связывания аммиака – синтез глутамина:
Локализация: митихондрии клеток
Фермент: глутаминсинтетаза (ингибируется АМФ, глюкозо-6-фосфатом, Гли, Ала и Гис)

Глутамин — основной донор азота в организме.

С током крови глутамин поступает в кишечник и почки.
Читайте также: Снимок мягких тканей ноги
В кишечнике под действием глутаминазы обр-ся глутамат, который подвергается
трансаминированию с пируватом – аминогруппа с глутамата переносится в состав аоланина, который поглощается печенью, а образовавшийся аммиак удаляется с фекалиями (5%), а другая часть выводится с почками (90%)


В почках также под действием глутаминазы происходит гидролиз глутамина
Ацидоз стимулирует активность глутаминазы почек для образования аммиака, который нейтрализует кислые продукты и в виде аммонийных солей экскретируется с мочой.
Эта реакция защищает организм от потери К и Na, которые аткже могут
использоваться для выведения анионов и утрачиваться
2. Еще одна реакция обезвреживания аммиака – синтез аспарагина

Под действием глутаминзависимой (животная ткань) и аммиакзависимой
(бактериальная клетка) аспарагинсинтетазы.
Этот путь используется редко, т.к. энергетически невыгоден.
Орнитиновый цикл мочевинообразования. Химизм, место протекания процесса. Энергетический эффект процесса, его регуляция. Количественное определение мочевины сыворотки крови, клиническое значение.
Синтез мочевины (орнитиновый цикл) – циклический процесс, состоит из 5
реакций, катализируемых 5 отдельными ферментами. Способствует выведению азота.
Место протекания: митохондрии и цитозоль
Транспорт аминного азота – в составе глутамина, аланина, аммиака

1. карбомоилфосфат-синтетаза 1




Механизм отрицательной обратной связи с участием аллостерических ферментов.
При уменьшении расхода АТФ – активность дыхательной цепи снижается (дыхательный контроль), конц. NADH в клетке повышается – снижение активности регуляторных ферментов.
NADH – ингибитор NAD-зависимые дегидрогеназы.
Повышение конц. цитрата, сукцинил-КоА и ацетил-КоА – ингибироваие регуляторных реакций.
Субстраты реакций – пируват и оксалоацетат – активируют ферменты ОПК
Биологическая роль орнитинового цикла:
Превращение азота АК в мочевину и предотвращение накопления аммиака
Синтез аргинина и пополнение его фонда в организме
Энергетический баланс процесса:
1 молекула мочевины за 1 оборот цикла расходует 4 макроэргические связи, которые компенсируются при дезаминировании глутамата и регенерации аспартата с образованием НАДН2
При трансмембранном переносе цитруллина и орнитина и переносе мочевины из крови в почки за счет градиента Na + , создаваемого К + , Na + -АТФазы.
Суммарное уравнение синтеза мочевины
СО2 + NH3 + аспартат + 3АТФ + 2Н2О = мочевина + фумарат +
+ 2 (АДФ + H3PO4) + АМФ + H4P2O7
Количественное определение мочевины сыворотки крови, клиническое значение
В опытную пробирку отмеривают в 10 раз сыворотки крови, в стандартную – такой
же объем стандартного раствора мочевины и в контрольную –дистиллированной воды.
Добавляют во все пробирки по 2 мл цветного реактива и тщательно перемешивают
их содержимое встряхиванием.
Все пробирки закрывают крышечкой из алюминиевой фольги и нагревают в
кипящей водяной бане в течение 10 мин, затем охлаждают их под струей холодной воды.
Измеряют оптическую плотность и опр. количественное содержание
по калибровочному графику.
Определение концентрации мочевины в крови применяется:
Для оценки тяжести патологического процесса,
Для наблюдения за течением заболевания
Оценка эффективности проводимого лечения.
Уровень мочевины в крови характеризует выделительную функцию почек
Декарбоксилирование аминокислот. Биогенные амины: гистамин, серотонин, ГАМК, путресцин. Реакции их образования, ферменты, кофактор. Биороль биогенных аминов. Дезаминирование и метилирование аминов как пути их обезвреживания.
В тканях млекопитающих декарбоксилированию может подвергаться целый ряд
аминокислот. Продуктами реакции являются СО2 и амины, которые оказывают выраженное биологическое действие на организм (биогенные амины)
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом