
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
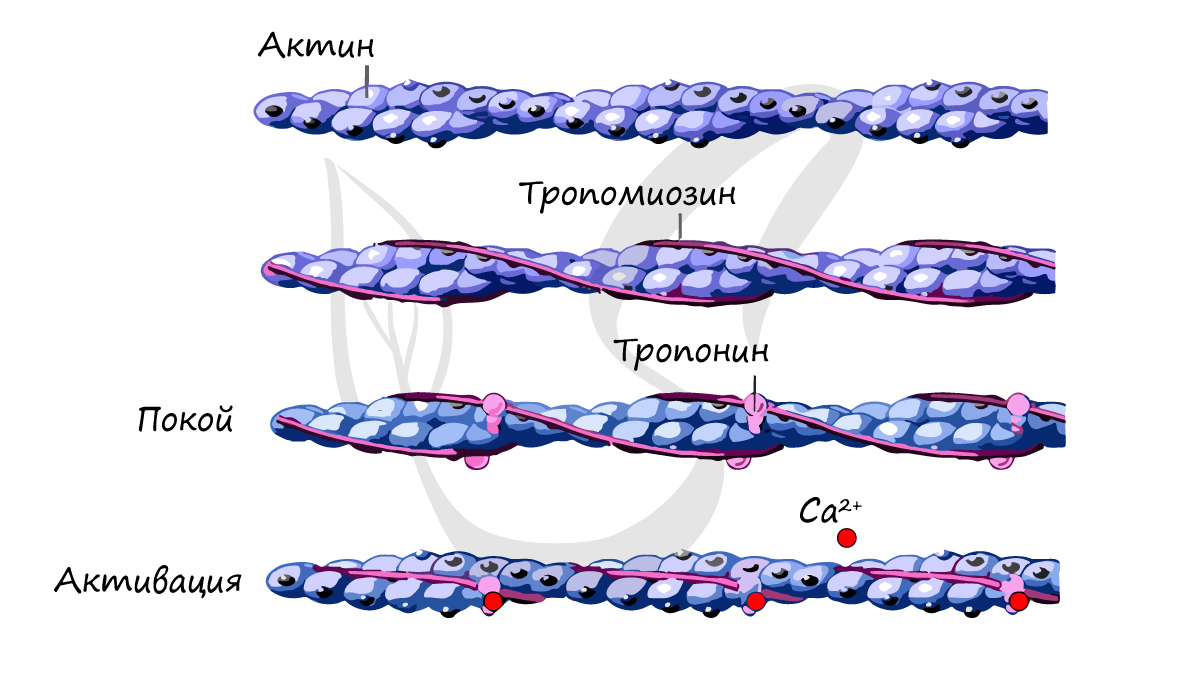
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Читайте также: Жировая ткань в разрезе
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Гладкие мышцы
Гладкие мышцы [ править | править код ]
В данной статье описаны основные характеристики гладкой мускулатуры. Рассматриваются следующие темы.
- Гладкие мышцы состоят из одиночных веретеновидных клеток.
- Как правило, гладкие мышцы выполняют непроизвольные сокращения.
- Гладкие мышцы являются важной составной частью стенок мышечных полых органов.
- В отличие от поперечно-полосатых мышц миофиламенты гладких мышц не имеют четкой организации и не имеют саркомеры.
- Гладкие мышцы можно тренировать с помощью специальных упражнений, чтобы повысить их эффективность и коэффициент полезного действия.
- Первичными контролирующими элементами гладкой мускулатуры в головном мозге являются нервные волокна вегетативной нервной системы.
- Гладкие мышцы являются основной составной частью стенок кровеносных и лимфатических сосудов.
- Тонус кровеносных сосудов определяет скорость и величину кровотока.
Гистологическое строение [ править | править код ]
В отличие от скелетных мышц, которые состоят из многоядерных элементов, образованных в результате слияния многих клеток, гладкая мышечная ткань образована отдельными клетками. Форма клеток приближается к веретеновидной (фузиформной), однако в отдельных случаях они могут иметь и другую форму (рис. 1.15). Клетки окружены базальной мембраной, содержащей большое количество белков, и имеют одно ядро в центре. В расслабленном состоянии клетки ядро продолговатое, а при сокращении принимает штопорообразную форму. При классическом окрашивании цитоплазма гладкомышечных клеток в световом микроскопе выглядит гомогенной. В гладкомышечной клетке отсутствуют саркомеры, поэтому ни в продольном, ни в поперечном срезе не видны миофибриллы (рис. 1.16). [[Image:|250px|thumb|right|рис. 1.16. Гистологическое строение гладких мышц — гладкомышечные веретенообразные клетки с одиночным расположенным в центре ядром; миофибриллы не видны]] Необходимые для сокращения актиновые и миозиновые филаменты прикрепляются либо к клеточной мембране (якорные бляшки), либо к так называемым плотным тельцам в цитоплазме.
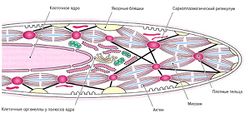
Ядро гладкомышечной клетки содержит хорошо выраженное ядрышко. Большинство клеточных органелл расположено около полюсов ядра. В отличие от скелетных мышц в гладкомышечных клетках слабо выражен саркоплазматический ретикулум, поэтому их сократимость значительно отличается (Welsch, 2006).
Формы и расположение гладких мышц [ править | править код ]
Гладкомышечные клетки помимо других свойств характеризуются непроизвольными сокращениями. Гладкие мышцы способны непроизвольно сокращаться, поэтому они не нуждаются в произвольном контроле. Соответственно этому, они расположены преимущественно в следующих органах человеческого организма.
- Гладкие мышцы являются важной составной частью стенок мышечных полых органов. К ним относится большинство органов пищеварительной и мочевыделительной систем, а также дыхательные пути.
- Пищеварительная система состоит из головной (полость рта и глотка) и туловищной частей пищеварительной трубки (пищевод, желудок и кишечник), а также связанных с ними экзокринных желез (слюнные железы, поджелудочная железа и печень).
- Железы содержат гладкие мышцы преимущественно в стенках их выводных протоков. Особенностью выводного протока печени является наличие разветвлений и расширения для депонирования секрета, которое называется желчным пузырем.
- Туловищная часть пищеварительного тракта (канала) имеет три слоя гладких мышц, ответственных за перистальтику и движение ворсинок слизистой оболочки кишки. В некоторых областях особенно выражена кольцевая мускулатура, которая образует сфинктеры или привратники и внутренний анальный сфинктер.
- В отличие от туловищной части головной конец пищеварительного тракта содержит поперечно-полосатые мышцы вместо гладких.
- Как и стенки пищеварительного тракта, стенки мочевыводящих путей также имеют три слоя гладкой мускулатуры, которые при более близком рассмотрении образуют единый спиралевидный слой.
- Как мужские, так и женские половые органы имеют большое количество гладких мышц.
- Гладкие мышцы в большом количестве содержатся в трахее и бронхах — важных частях нижних дыхательных путей. Гладкомышечные клетки регулируют объем поступающего воздуха в зависимости от внешних факторов за счет изменения диаметра дыхательных путей.
Гладкие мышцы являются составной частью кровеносных и лимфатических сосудов. Количество гладкомышечной ткани пропорционально диаметру сосуда, причем при одинаковых размерах артерии содержат больше гладкой мускулатуры, чем вены и лимфатические сосуды. По мере разветвления сосудов уменьшается и абсолютное количество гладких мышц, при этом в артериолах относительно их диаметра содержание гладкой мышечной ткани максимально. В капиллярах гладкомышечная ткань отсутствует.
Гладкомышечная ткань также встречается в некоторых немышечных органах.
- В глазном яблоке гладкие мышцы ответственны за расширение и сужение зрачка, а также натяжение капсулы хрусталика. Это позволяет изменять его преломляющую силу и количество света, падающего на сетчатку. Этот механизм важен для ближнего и дальнего зрения и восприятия глубины.
- Поднимание волос как реакция на холод или страх выполняется мышцами, поднимающими волос (шш. arrectores pilorum). При их сокращении кожа приобретает вид «гусиной кожи» («мурашки»). Также в организме (например, в экзокринных железах или яичнике при овуляции) встречаются клетки, представляющие собой нечто среднее между мышечными и соединительнотканными клетками (так называемые миофибробласты) или клетками эпителия (миоэпителиоциты) (Drenckhahn, 2003).

Запомните:Необходимо упомянуть, что некоторые мышцы, выполняющие более или менее непроизвольные сокращения, являются поперечно-полосатыми. К ним относятся следующие мышцы.
- Диафрагма — главная дыхательная мышца.
- Сердечная мышца.
- Мышцы дна ротовой полости, нёба и глотки.
- Мышцы гортани.
- Мимические мышцы.
- Мышцы тазового дна и наружные сфинктеры (анальный и сфинктер уретры).
Строение гладких мышц [ править | править код ]
Как и у поперечно-полосатых мышц, сокращение гладких мышц происходит за счет взаимного скольжения миофиламентов — тонких актиновых и толстых миозиновых филаментов (миозин II типа). Гладкомышечные клетки содержат в 3 раза меньше миозина, чем поперечно-полосатые. В отличие от скелетных мышц миозиновые филаменты в гладкомышечных клетках расположены неупорядоченно и не образуют саркомеры и миофибриллы, что и послужило причиной появления термина «гладкие мышцы». Тонкие актиновые филаменты, как уже было указано, прикрепляются к плотным тельцам в цитоплазме или к якорным бляшкам клеточной мембраны. Эти образования являются аналогами Z-линии в поперечно-полосатых мышцах. Помимо хаотичного расположения миофиламентов сократительный аппарат гладких мышц отличается от поперечно-полосатых как ультраструктурно, так и биохимически. Одним из важных структурных различий является активность Са2+-каналов
Читайте также: Для чего служит покровная ткань у растений
и фермента АТФазы миозина, которая влияет на скорость мышечных сокращений. Плохо развитый саркоплазматический ретикулум позволяет депонировать лишь небольшое количество ионов Са2+, а большая часть ионов, необходимых для мышечного сокращения, поступает при возбуждении клетки из межклеточного пространства. По этой причине в гладкомышечной ткани наблюдается, с одной стороны, медленный ток ионов Са2+, а с другой — меньшая активность АТФазы миозина (в 10-100 раз ниже, чем в поперечно-полосатых мышцах). Таким образом, сокращения гладких мышц характеризуются низкой скоростью, но большей длительностью (Widmaier et al., 2008).
Сократительная способность [ править | править код ]
Характер, процесс и контроль мышечных сокращений в гладких мышцах значительно отличается от таковых в поперечно-полосатых мышцах. Последние способны к быстрым сокращениям и быстро утомляются, в то время как гладкие мышцы характеризуются относительно медленными сокращениями, но обладают большей выносливостью. Причиной этого является особая ультраструктура гладкомышечных клеток, а также молекулярная структура миофиламентов. Кроме этого, деполяризация клеточной мембраны (и, следовательно, поступление ионов Са2+ и возникающее сокращение) гладкомышечных клеток вызывается множеством факторов, в то время как деполяризация мембраны скелетных мышц регулируется нейромедиатором ацетилхолином (АЦХ), выделяемым двигательными нейронами.
Именно последняя особенность считается причиной непроизвольности сокращений гладких мышц. Иногда утверждают, что гладкие мышцы вообще не способны к произвольным сокращениям, однако это не всегда так. Более правильно утверждать, что гладкие мышцы не нуждаются в произвольном контроле, т. к. центры в стволе головного мозга, ответственные за кровообращение, функцию пищеварительного тракта и т. д., функционируют без сознательного контроля. Это также справедливо для нервных центров, регулирующих функцию сердца и дыхания, хотя сердечная мышца и дыхательные мышцы (особенно диафрагма) являются поперечно-полосатыми.
Необходимо отметить, что контроль скелетных мышц только частично произволен. Единственные мышцы, которые поодиночке выполняют истинные произвольные движения (причем некоторые только при тренировке), — мышцы кисти. Все остальные движения возникают в результате сложных неосознанных взаимодействий многих мышц (мышц-синергистов), т. к. при этом всегда изменяются статические силы тела. Для простого сгибания руки в локте требуется напряжение сгибателей кисти, лучезапястного сустава и локтевого сустава. При этом должны также одновременно расслабиться мышцы-антагонисты (все разгибатели соответствующих суставов, например трехглавая мышца плеча). Дополнительно активируются различные мышцы плечевого пояса, которые стабилизируют лопатку относительно туловища, а также различные мышцы, стабилизирующие туловище, тазобедренный сустав и нижние конечности, чтобы поддерживать статику (позу) тела. Из этого примера видно, что произвольное напряжение одной мышцы, не говоря уже об отдельных мышечных волокнах, невозможно, как и произвольное напряжение отдельных гладких мышц. Возможна лишь активация мышечной системы, а именно совокупности синергистов для выполнения одного движения (например, сгибания локтевого сустава).
Учитывая вышесказанное, читателя не должно удивить, что произвольная активация системы гладких мышц все же возможна. К примеру, с помощью методов биологической обратной связи или расслабления в результате тренировок возможно относительно легко получить контроль над активностью мышц сердечно-сосудистой системы. Эти методы часто применяются в лечении мигреней и артериальной гипертензии.
Кроме этого, было продемонстрировано, что различный уровень тренированности влияет не только на внутри- и межмышечную координацию скелетных мышц, но и на активность гладких мышц сердечнососудистой системы (артериальное давление) или желез внешней секреции (выделение пота), которая может значительно изменяться под влиянием спортивных тренировок.
В заключение необходимо отметить, что гладкие мышцы до определенного предела все же подчиняются произвольному контролю. Из этого следует очевидный вывод, который давно известен в спорте: гладкие мышцы можно тренировать с помощью специальных методик и повышать их работоспособность и эффективность. Простые тренировки на выносливость через несколько недель позволяют значительно повысить эффективность работы, например, сердечно-сосудистой системы. Схожие результаты известны и для потовых желез: после упражнений на выносливость тренированные люди начинают потеть раньше, чем нетренированные.
Физиология сокращения гладких мышц [ править | править код ]
Отличия физиологических функций гладких мышц определяют их биохимические и анатомические особенности. Наиболее важными являются следующие особенности (Widmaier et al., 2008).
- Скорость сокращений — сокращение гладкомышечных клеток, подобно остальных типам мышечной ткани, основано на повышении концентрации ионов Са2+ в цитоплазме. В отличие от поперечно-полосатых мышц в них слабо развит саркоплазматический ретикулум, поэтому они становятся зависимы от поступления ионов в клетку. Входящий ток ионов значительно медленнее, чем процесс высвобождения Са2+ из саркоплазматического ретикулума, кроме того, активность ферментов миозина в гладкомышечных клетках в 10-100 раз ниже, чем в поперечно-полосатых. Эти два фактора обусловливают относительно низкую скорость сокращений гладких мышц.
- Выносливость — расслабление гладкой мускулатуры происходит после разрыва поперечных связей между актиновыми и миозиновыми филаментами. Для этого необходимо дефосфорилирование головки миозина, т. к. он способен связываться с актином только в фосфорилированном состоянии. При возбуждении гладкомышечной клетки постоянно происходят процессы фосфорилирования и дефосфорилирования миозиновых головок, причем скорость фосфорилирования выше, чем скорость дефосфорилирования. При длительном повышении концентрации Са2+ активируются процессы дефосфорилирования головок миозина, уже связанных с актином. Таким образом, гладкомышечные клетки могут в течение многих часов без выраженного утомления и потребления большого количества энергии поддерживать статическое сокращение. Это можно сравнить с трупным окоченением скелетных мышц, однако это происходит в живом организме и типично для гладкомышечных сфинктеров (например, выходной сфинктер мочевого пузыря).
- Физиологическая недостаточность — благодаря своей ультраструктуре гладкие мышцы могут совершать сильные изометрические и концентрические сокращения в большем диапазоне длины, чем скелетные мышцы. В качестве примера можно привести сильное растяжение мочевого пузыря или матки в конце беременности (в этих случаях мышцы могут растягиваться в 8 раз).
Контроль сокращений [ править | править код ]
В отличие от скелетных мышц гладкие и сердечные мышцы способны к спонтанной деполяризации и сокращению, независимому от нервных стимулов. В сердце процессы спонтанной деполяризации являются одной из функций органа, имеют систему внешнего контроля и выполняются в здоровом сердце только группой специфических мышечных клеток (клетки синусного и атриовентрикулярного узлов). В гладких мышцах существует множество факторов, которые могут вызывать эти процессы и на них влиять.
Вегетативная нервная система [ править | править код ]
Первичным органом контроля гладких мышц являются центры вегетативной нервной системы (ВНС, автономная нервная система). Филогенетически это очень старая часть нервной системы, расположенная преимущественно в стволе головного мозга и контролируемая гипоталамусом. ВНС участвует в поддержании важных параметров гомеостаза и адаптации к изменяющимся условиям среды, например к физической нагрузке: повышение тонуса сосудов, расширение дыхательных путей, уменьшение перистальтики кишечника и т. д. без участия непосредственного контроля со стороны коры больших полушарий. Существует три типа ВНС.
- Внутрикишечная, или энтерическая, вегетативная нервная система (ЭНС — энтеральная, или интрамуральная, нервная система) — скопление нервных клеток в стенке кишечника. Как и некоторые гладкомышечные или сердечные клетки, они обладают способностью к спонтанному генерированию потенциалов действия и вызывают ритмическое волнообразное сокращение мышц кишечника (перистальтику). Предположительно, ЭНС — единственная часть нервной системы, обладающая истинной автономностью и не требующая контроля головного мозга, чья активность модулируется только другими центрами ВНС. Перистальтика кишечника продолжается даже после удаления его из организма до полного исчерпания запаса электролитов, кислорода и питательных веществ.
- Симпатическая нервная система — общей функцией данной системы является повышение активности внутренних органов, необходимых при стрессовых ситуациях (борьба, бегство, сексуальная активность и спорт). Симпатическая нервная система повышает активность сердца, тонус кровеносных сосудов и артериальное давление, расширяет верхние дыхательные пути и, следовательно, увеличивает поступление кислорода, расширяет зрачки, усиливает потоотделение и т. д. Одновременно с этим она уменьшает активность кишечника, продукцию мочи и органов выделения. Веществами, через которые симпатическая нервная система оказывает влияние на гладкие мышцы, в первую очередь считаются нейромедиаторы адреналин и норадреналин (приблизительно в соотношении 80:20).
- Парасимпатическая нервная система — рассматривается как антагонист симпатической нервной системы, т. к. она оказывает противоположное действие. В остальном же взаимодействие между двумя типами рассматривается как синергическое, т. к. задачей парасимпатической нервной системы является накопление энергии, требуемой симпатической системе. Парасимпатическое влияние на внутренние органы преобладает в состоянии покоя и реактивной фазе сна. Синергическое действие обеих систем также видно на примере процесса полового акта, т. к. эрекция — процесс, контролируемый парасимпатической системой, а эякуляция — симпатической. Нейромедиатором парасимпатической системы является ацетилхолин (АЦХ).
Читайте также: Абажур для торшера своими руками из ткани выкройки
Помимо вышеперечисленных нейромедиаторов на сократимость гладких мышц влияет множество веществ, например моноокись азота и серотонин или яды растительного происхождения мускарин (мухоморы), атропин (красавка/белладонна), никотин (табачные растения) и кураре (южноамериканская лилия).
Высвобождение Са2+ при растяжении [ править | править код ]
Потенциалы действия в гладких мышцах также возникают при их растяжении. При этом открываются чувствительные к растяжению Са2+-каналы в клеточной мембране и ионы Са2+ устремляются в клетку. Данный механизм не только чрезвычайно важен для поддержания гомеостаза, но и является причиной возникновения колик. При этом гладкие мышцы полых органов (мочеточников, желчевыводящих путей, кишечника и т.д.) растягиваются конкрементом, что приводит к их рефлекторному сокращению. Присутствие конкремента нарушает моторику этих органов, сокращение мышц не позволяет продвинуть конкремент, что вызывает новые сокращения (периодические спазмы) (Widmaier et al., 2008).
Физиология гладких мышц [ править | править код ]
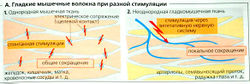
Гладкая мышца состоит из многих слоев веретеновидных клеток. Гладкие мышцы работают во многих органах (желудок, кишечник, желчный пузырь, мочевой пузырь, матка, бронхи, глаз и т. д.), а также в кровеносных сосудах, где играют важную роль в регуляции кровообращения. Гладкие мышцы содержат специальный тип F-актин-тропомиозин- и миозин II-филаментов, но мало тропонина и миофибрилл. Кроме того, гладкие мышцы не имеют развитой системы микротрубочек и саркомеров (они не исчерчены). Отсюда и название — гладкая мускулатура. Гладкомышечные филаменты формируют слабый сократительный аппарат, расположенный в клетке продольно и прикрепленный к дисковидным бляшкам (модель см. Б), которые также обеспечивают механическую связь между клетками в гладкой мускулатуре. Гладкая мышца может укорачиваться гораздо сильнее, чем поперечно-полосатая мышца.
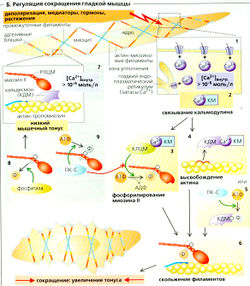
Мембранный потенциал гладкомышечных клеток многих органов (например, кишечника) не постоянный, а ритмично изменяется с низкой частотой (от 3 до 15 мин-1) и амплитудой (от 10 до 20 мВ), таким образом образуя медленные волны. Эти волны вызывают вспышки потенциала действия (пики), если превосходят некоторый потенциал покоя. Чем дольше медленная волна остается выше потенциала покоя, тем больше количество и частота потенциалов действия, которые она производит. Сравнительно вялое сокращение происходит примерно через 150 мс после пика. Тетанус наступает при довольно низкой частоте пиков. Следовательно, гладкая мускулатура постоянно находится в состоянии более или менее сильного сокращения (тонуса). Потенциал действия гладкомышечных клеток некоторых органов имеет плато, аналогичное таковому у сердечного потенциала действия.
Существуют два типа гладкой мускулатуры (А). Клетки однородной гладкомышечной ткани электрически сопряжены друг с другом щелевыми контактами. В таких органах, как желудок, кишечник, желчный пузырь, мочевой пузырь, мочеточники, матка и в некоторых типах кровеносных сосудов, т. е. там, где присутствует этот тип гладкомышечной ткани, стимулы передаются от клетки к клетке. Стимулы генерируются автономно изнутри гладкой мускулатуры (частично клетками-водителями ритма). Другими словами, стимул не зависит от иннервации и во многих случаях является спонтанным (миогенный тонус). Второй тип, неоднородная гладкомышечная ткань, характеризуется тем, что межклеточные взаимодействия там осуществляются в основном при помощи стимулов, передаваемых вегетативной нервной системой (нейрогенный тонус). Это происходит в таких структурах, как артериолы, семенные канальцы, радужная оболочка глаза, ресничное тело, мышцы у корней волос. Поскольку между этими гладкомышечными клетками обычно нет щелевых контактов, стимуляция остается локальной, как в двигательных единицах скелетной мышцы.
Тонус гладкой мускулатуры регулируется степенью деполяризации (например, посредством растяжения или клеток — водителей ритма), а также с помощью медиаторов (например, ацетилхолина или норадреналина) и многочисленных гормонов (например, в матке — эстрогена, прогестерона и окситоцина, в стенках сосудов — гистамина, ангиотензина II, вазопрессина, серотонина и брадикинина). Увеличение тонуса происходит, если любой из этих факторов прямо или опосредованно увеличивает внутриклеточную концентрацию Са 2+ до более 10-6 моль/л. Приток Са 2+ происходит в основном из межклеточного пространства, но небольшие количества поступают и из внутриклеточных запасов (Б1). Ионы Са 2+ связываются кальмодулином (КМ) (Б2), и комплекс Са 2+ -КМ способствует сокращению. Как же это происходит?
Регуляция на уровне миозина II (БЗ): комплекс Са 2+ -КМ активирует киназу легких цепей миозина (КЛЦМ), которая фосфорилирует регуляторную легкую цепь миозина (РЛЦМ) по заданному положению, благодаря чему миозиновая головка может взаимодействовать с актином (Б6).
Регуляция на уровне актина (Б4): комплекс Са 2+ -КМ также связывает кальдесмон (КДМ), который затем отщепляется от комплекса актин-тропомиозин, делая его доступным для скольжения филаментов (Б6). Фосфорилирование КДМ протеинкиназой С (ПК-С) также, по-видимому, способно индуцировать скольжение филаментов (В5).
Следующие факторы ведут к снижению тонуса: снижение концентрации Са 2+ в цитоплазме ниже 10-6 моль/л (Б7), фосфатазная активность (Б8), а также активность протеинкиназы С, когда она фосфорилирует другое положение легкой цепи миозина (Б9).
При регистрации зависимости «длина-сила» для гладкой мышцы выявляется постоянное снижение мышечной силы, в то время как длина мышцы остается постоянной. Это свойство мышцы называется пластичностью.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом