
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
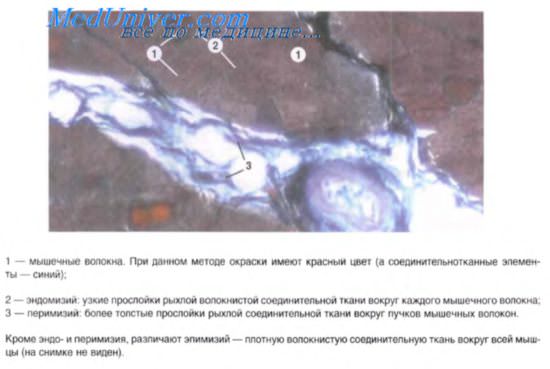
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Читайте также: Бусы из цветов из ткани своими руками
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
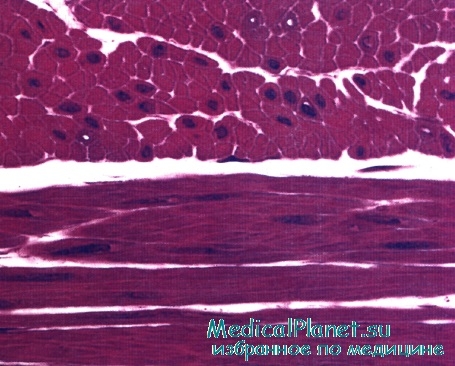
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
Читайте также: Ткань для шубки тедди
Гладкая мышечная ткань желудка функции
Строение желудка. Стенки желудка. Мышцы желудка. Слизистая желудка.
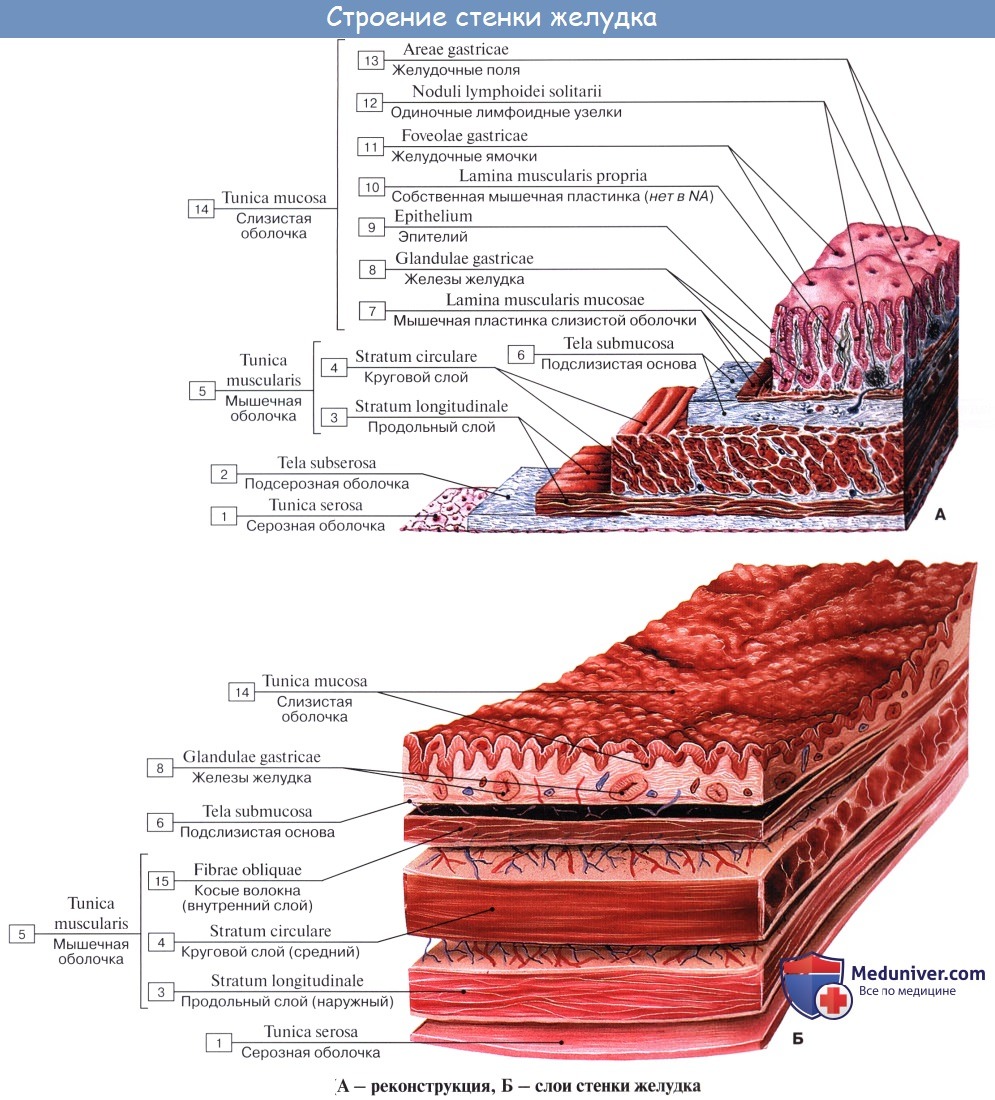
Стенка желудка состоит из трех оболочек:
1) tunica mucosa — слизистая оболочка с сильно развитой подслизистой основой, tela submucosa;
2) tunica muscularis — мышечная оболочка;
3) tunica serosa — серозная оболочка.
Tunica mucosa построена соответственно основной функции желудка — химической обработке пищи в условиях кислой среды.
В связи с этим в слизистой имеются специальные желудочные железы, вырабатывающие желудочный сок, succus gastricus, содержащий соляную кислоту.
Различают три вида желез:
1) кардиальные железы, glandulae cardiacae;
2) желудочные железы, glandulae gastricae (propriae); они многочисленны (приблизительно 100 на 1 мм 2 поверхности), расположены в области свода и тела желудка и содержат двоякого рода клетки: главные (выделяют пепсиноген) и обкладочные (выделяют соляную кислоту);
3) пилорические железы, glandulae pyloricae, состоят только из главных клеток.
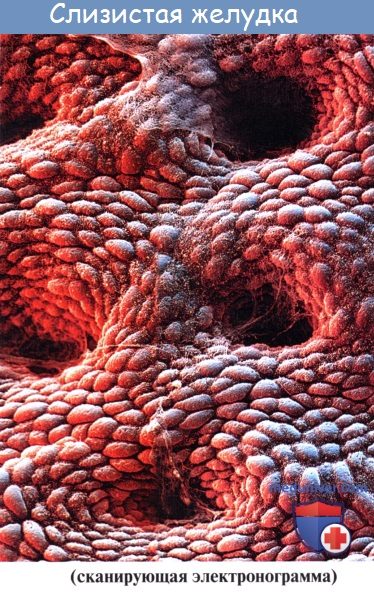
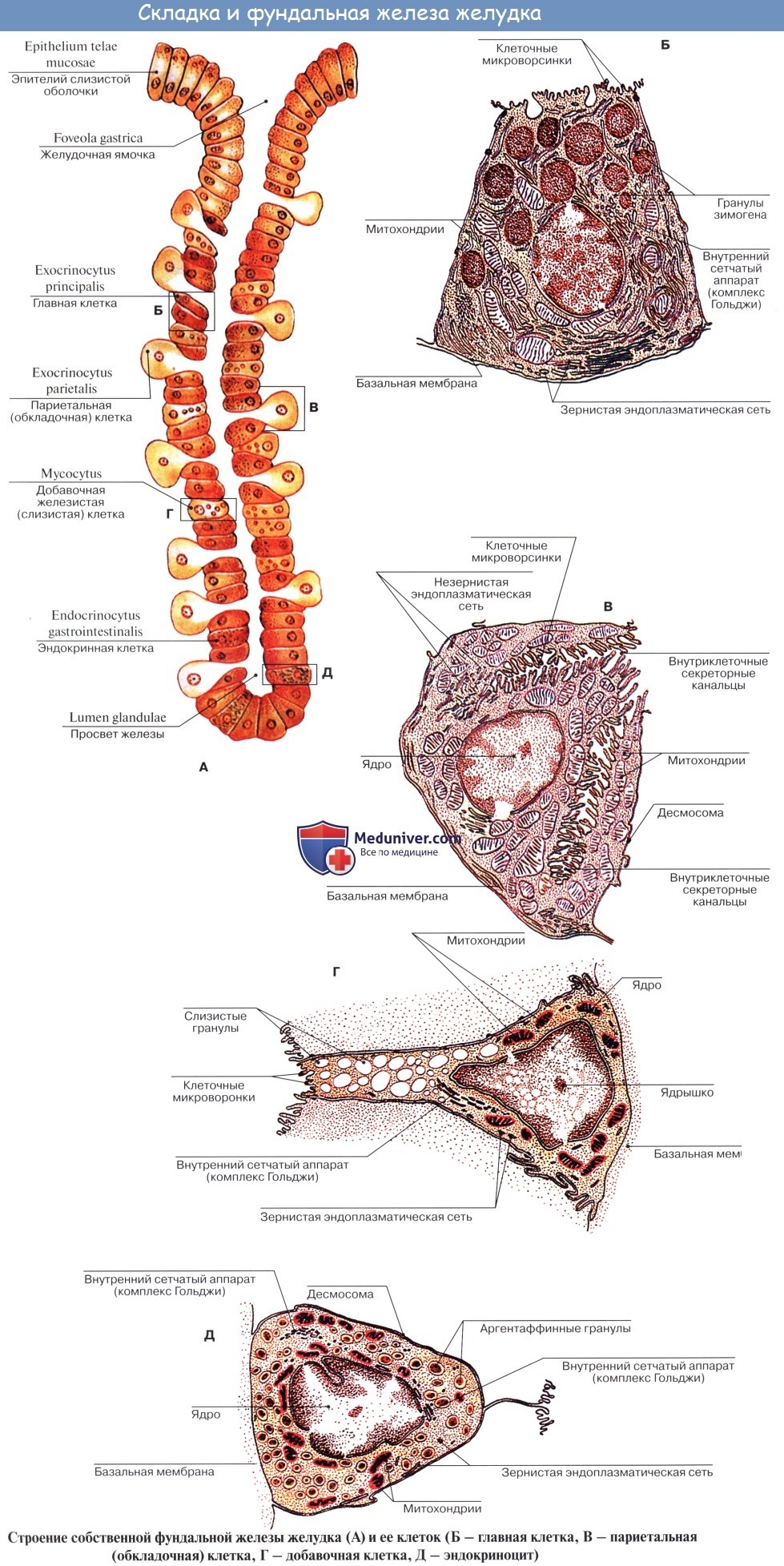
Местами в слизистой разбросаны одиночные лимфатические фолликулы, folliculi lymphatici gastrici.
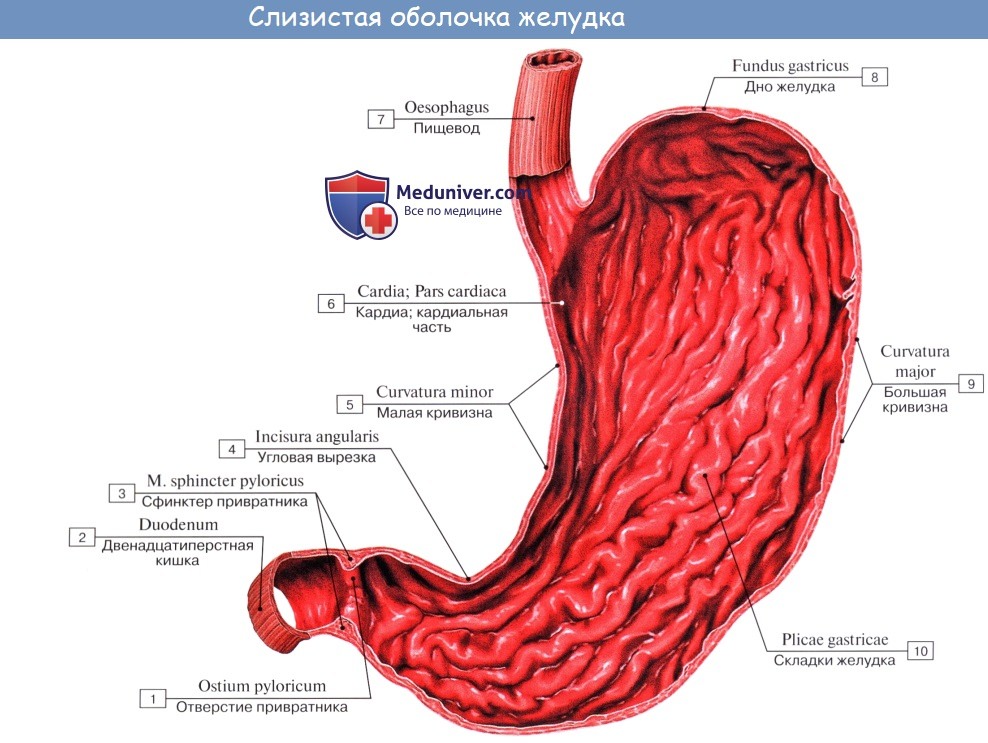
Тесное соприкосновение пищи со слизистой оболочкой и лучшее пропитывание ее желудочным соком достигается благодаря способности слизистой собираться в складки, plicae gastricae, что обеспечивается сокращением собственной мускулатуры слизистой (lamina muscularis mucosae) и наличием рыхлой подслизистой основы, tela submucosa, содержащей сосуды и нервы и позволяющей слизистой оболочке сглаживаться и собираться в складки различного направления.
Вдоль малой кривизны складки имеют продольное направление и образуют «желудочную дорожку», которая при сокращении мышц желудка может стать в данный момент каналом, по которому жидкие части пищи (вода, солевые растворы) могут проходить из пищевода в привратник, минуя кардиальную часть желудка.
Кроме складок, слизистая имеет кругловатые возвышения (диаметром 1 — 6 мм), называемые желудочными полями, areae gastricae, на поверхности которых видны многочисленные маленькие (0,2 мм в диаметре) отверстия желудочных ямок, foveolae gastricae. В эти ямки и открываются железы желудка.
В свежем состоянии tunica mucosa красновато-серого цвета, причем на месте входа пищевода макроскопически заметна резкая граница между плоским эпителием пищевода (эпителием кожного типа) и цилиндрическим эпителием желудка (эпителием кишечного типа).
В области отверстия привратника, ostium pyloricum, располагается циркулярная складка слизистой оболочки, отграничивающая кислую среду желудка от щелочной среды кишечника; она называется valvula pylorica.
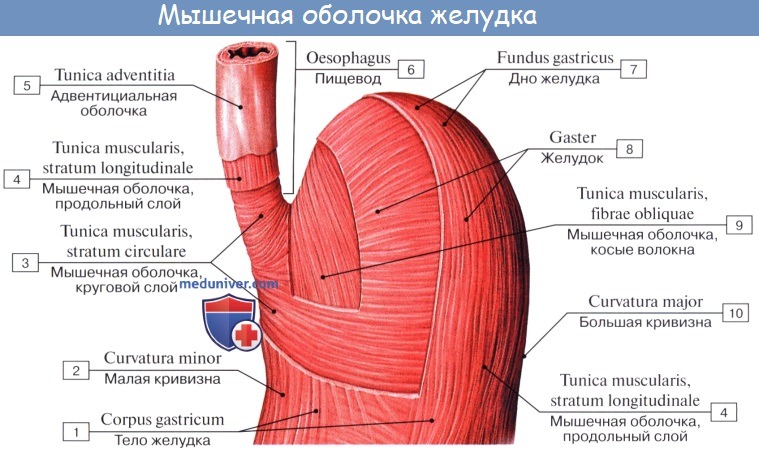
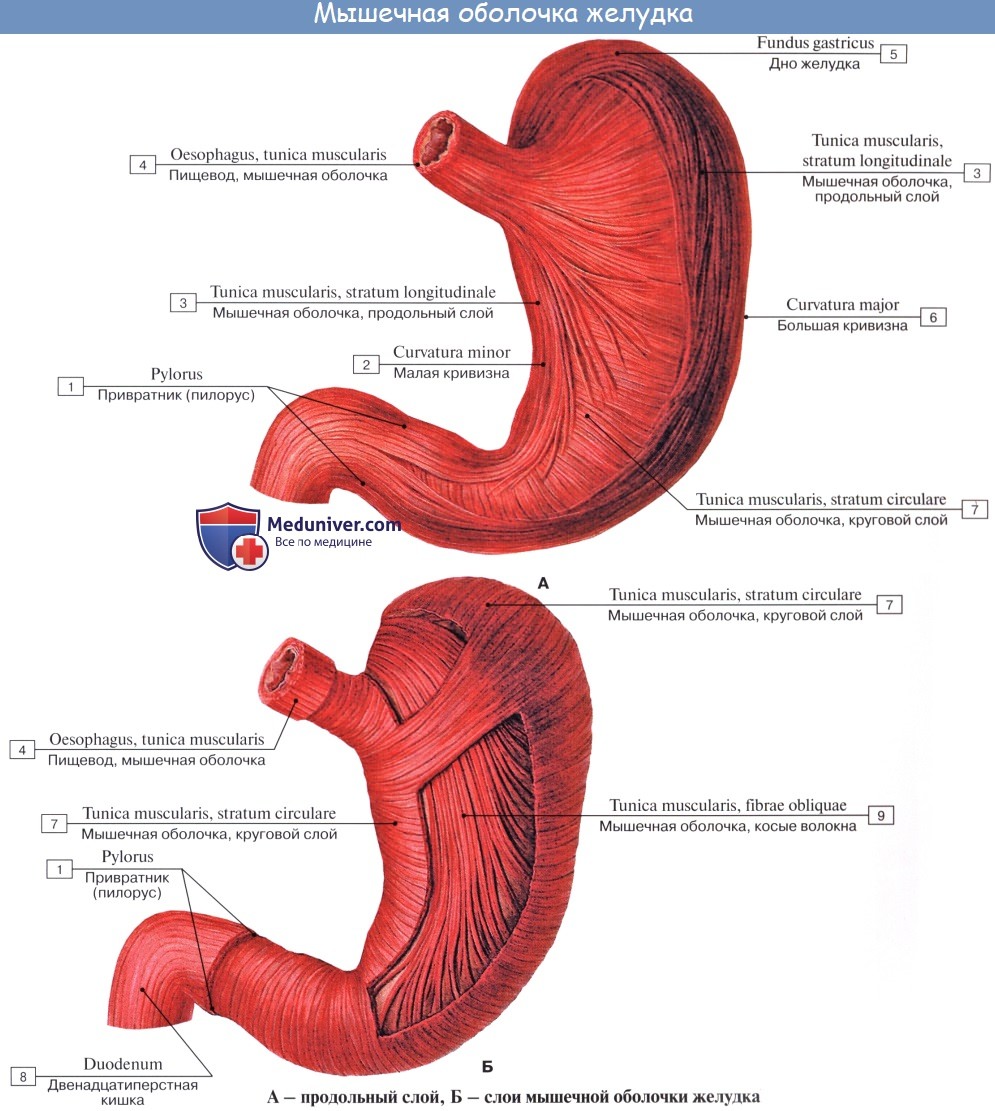
Tunica muscularis представлена миоцитами, неисчерченной мышечной тканью, которые способствуют перемешиванию и продвижению пищи; соответственно форме желудка в виде мешка они располагаются не в два слоя, как в пищеводной трубке, а в три: наружный — продольный, stratum longitudindle; средний — циркулярный, stratum circulare, и внутренний — косой, fibrae obliquae. Продольные волокна являются продолжением таких же волокон пищевода.
Stratum circularе выражен сильнее продольного; он является продолжением циркулярных волокон пищевода. По направлению к выходу желудка циркулярный слой утолщается и на границе между pylorus и двенадцатиперстной кишкой образует кольцо мышечной ткани, m. sphincter pylori — сжиматель привратника.
Соответствующая сфинктеру привратниковая заслонка, valvula pylorica, при сокращении сжимателя привратника совершенно отделяет полость желудка от полости двенадцатиперстной кишки.
Sphincter pylori и valvula pylorica составляют специальное приспособление, регулирующее переход пищи из желудка в кишку и препятствующее обратному ее затеканию, что влекло бы за собой нейтрализацию кислой среды желудка.
Fibrae obliquae, косые мышечные волокна, складываются в пучки, которые, охватывая петлеобразно слева ostium cardiacum, образуют «опорную петлю», служащую punctum fixum для косых мышц. Последние спускаются косо по передней и задней поверхностям желудка и при своем сокращении подтягивают большую кривизну по направлению к ostium cardiacum.
Самый наружный слой стенки желудка образуется серозной оболочкой, tunica serosa, которая представляет собой часть брюшины; серозный покров тесно срастается с желудком на всем его протяжении, за исключением обеих кривизн, где между двумя листками брюшины проходят крупные кровеносные сосуды.
На задней поверхности желудка влево от ostium cardiacum имеется небольшой участок, не прикрытый брюшиной (около 5 см ширины), где желудок непосредственно соприкасается с диафрагмой, а иногда с верхним полюсом левой почки и надпочечником. Несмотря на свою сравнительно простую форму, желудок человека, управляемый сложным иннервационным аппаратом, является весьма совершенным органом, позволяющим человеку довольно легко приспособляться к различным пищевым режимам.
Ввиду легкого наступления посмертных изменений формы желудка и невозможности поэтому результаты наблюдений на трупе целиком переносить на живого, большое значение получает исследование с помощью гастроскопии и особенно рентгеновских лучей.
Учебное видео по анатомии глотки, пищевода, желудка
Редактор: Искандер Милевски. Дата последнего обновления публикации: 23.07.2021