
Из протекающей артериальной крови ткани усваивают 6 кислорода
Д-р. Роб Лоу, Королевская больница, Бристоль, Великобритания
Д-р. Х. Буквирва, больница Мулаго, Кампала, Уганда
Для выживания человек должен быть способен поглощать кислород из атмосферы и транспортировать его клеткам, где он используется в метаболизме. Некоторые клетки могут короткое время вырабатывать небольшое количество энергии без участия кислорода (анаэробный метаболизм). Другие органы (например, головной мозг) состоят из клеток, которые могут существовать только при наличии постоянного снабжения кислородом (аэробный метаболизм). Различные ткани имеют различную степень толерантности к аноксии (отсутствие кислорода). Мозг и сердце – наиболее уязвимые органы. В начале недостаток кислорода поражает функцию органа, а с течением времени вызывает и необратимые морфологические изменения (втечение минут в случае с мозгом), когда восстановление функции невозможно.
Транспорт кислорода из воздуха тканям
Кислород доставляется из воздуха, которым мы дышим, каждой клетке организма. В целом, газы перемещаются из области высокой концентрации (давления) в зону низкой концентрации (давления). Если в резервуаре имеется смесь газов, то давление каждого газа (парциальное давление) равняется давлению, при котором находился бы газ, будь он в резервуаре один.
Воздух (атмосфера) вокруг нас имеет давление 760 мм Hg (1 атмосфера = 760 мм Hg = 101 кРа). Воздух содержит 21% кислорода, 78% азота и небольшое количество СО2, аргона и гелия. Давление, создаваемое двумя основными газами, смешанными вместе, равняется общему или атмосферному давлению. Давление кислорода (РО2) в сухом воздухе на уровне моря равняется 159 мм Hg (21/100 х 760 = 159). При вдохе воздух, продвигающийся по верхним дыхательным путям, нагревается и увлажняется. Влажность, формирующаяся парами воды, создает давление, которое при 37 градусах в трахее составляет 47 мм Hg. Беря в расчет давление водяных паров, РО2 в трахее при дыхании воздухом (760-47) х 21/100 = 150 мм Hg. При достижении альвеол РО2 падает до 100 мм Hg. Это происходит из-за того, что в альвеолах идет процесс постоянного поступления О2 извне (дыхание) и удаления его из организма легочными капиллярами.
Кровь, возвращающаяся к сердцу из тканей, имеет низкое РО2 (40 мм Hg). Она идет к легким по легочным артериям. Легочные артерии образуют легочные капилляры, окружающие альвеолы. Кислород диффундирует из альвеол – зоны высокого РАО2 (100 мм Hg) в капилляры – зону низкого РаО2 (40 мм Hg). После оксигенации кровь поступает по легочным венам в левые отделы сердца и далее в ткани организма. В «идеальном легком» РаО2 крови в легочных венах должно равняться РАО2 в альвеолах. Три фактора делают РО2 венозной крови ниже альвеолярного: нарушение вентиляционно/перфузионных (V/Q) отношений, шунт и медленная диффузия.
Нарушение вентиляционно/перфузионных отношений
В «идеальном легком» все альвеолы получали бы одинаковую степень вентиляции и капилляризации с одинаковым током крови по ним.
Пораженные легкие могут иметь значительный дисбаланс между вентиляцией и кровотоком. Некоторые альвеолы сравнительно гипервентилируются, другие же «гиперперфузируются». Наиболее яркой формой этого нарушения является шунт, когда кровь протекает по альвеолам без газообмена (рисунок 1). Хорошо вентилируемые альвеолы (с высоким РО2 в капиллярной крови) не способны компенсировать газообмен при наличии большого числа невентилируемых альвеол с низким РО2 в капиллярах. Это происходит из-за того, что гемоглобин способен нести лимитированное количество кислорода (см. кривую диссоциации оксигемоглобина, рис. 2а). Легочная венозная кровь (смешанная кровь всех легочных капилляров) будет иметь меньшее РО2, чем в альвеолах. Даже нормальные легкие имеют некоторый вентиляционно/перфузионный дисбаланс; верхние зоны сравнительно лучше вентилируются, тогда как нижние – лучше перфузируются, но гиповентилируются.
Рис. 1. Отражена норма и два типа V/Q нарушений, таких как шунт и мертвое пространство.
Шунт имеет место, когда деоксигенированная венозная кровь, возвращающаяся от тканей проходит через невентилируемые альвеолы и поступает в легочные вены и большой круг кровообращения с неизмененным РО2 (40 мм Hg) (рисунок 1). Ателектазы (коллабированные альвеолы), уплотнение легкого, отек легких или обструкция дыхательных путей малого калибра (см. ниже) вызывают развитие шунта.
Кислород диффундирует из альвеол в капилляры до момента выравнивания РО2 в капиллярах и альвеолах. Этот процесс завершается, когда кровь проходит одну треть пути по легочному капилляру. В интактных легких процесс диффузии происходит очень быстро и завершается даже при высоком сердечном выбросе (нагрузка), когда кровь находится меньшее время в контакте с альвеолами. Этого может не происходить в поврежденной капиллярной сети легких (легочные заболевания). Однако, легкие имеют широкий диапазон компенсации и проблемы, связанные с неадекватной диффузией газов – редкая причина гипоксии, за исключением случаев альвеолярного фиброза.
Для нивелирования негативных эффектов легочного шунта и вентиляционно/перфузионных нарушений на оксигенацию крови легочные сосуды обладают способностью к вазоконстрикции — таким образом снижается кровоток по плохо вентилируемым зонам. Это называется гипоксической легочной вазоконстрикцией, снижающей эффект шунта.
Транспорт кислорода кровью
Кислород переносится кровью в двух формах. Большая часть связывается с гемоглобином (рисунок 2b), но также имеется очень небольшая доля кислорода, растворенного в плазме. Каждый грамм гемоглобина при полном насыщении способен переносить 1,31 мл кислорода. Таким образом, каждый литр крови с концентрацией гемоглобина 15 г/дл (150 г/л – пер.) может переносить около 200 мл кислорода при полном насыщении (РО2 >100 мм Hg). При этом РО2 всего 3 мл кислорода растворится в каждом литре плазмы.
Читайте также: Платье из белого с черным ткани
Если РаО2 значительно повышено (при дыхании 100% кислородом), небольшое количество кислорода растворится в плазме (0,003 мл О2/100 мл крови/мм Hg РО2), но при этом гемоглобин при сатурации > 95% не способен продолжать связываться с кислородом. При рассмотрении адекватности доставки кислорода тканям следует иметь в виду три фактора: концентрация гемоглобина, сердечный выброс и оксигенация.
Кислород перемещается по градиенту из зоны более высокой концентрации в воздухе через дыхательные пути, смешиваясь с альвеолярным газом, далее поступает в артериальную кровь, капилляры и, наконец, в клетку. РО2 достигает минимального уровня (4-20 мм Hg) в митохондриях (структурное образование клетки, ответственное за продукцию энергии). Это снижение парциального давления кислорода от воздуха к митохондрии известно как кислородный каскад, каждая ступень которого может быть затронута патологическим процессом и вызывать гипоксию (рисунок 3).
Рис. 2. Кривая диссоциации HbО2 при рН=7,4, РСО2= 40 ммHg и 37оС. Рис. 2b. Относительные доли О2, растворенного в плазме, и О2, связанного с Hb (концентрация Hb = 15г/100 мл крови).
Количество кислорода, доступного для организма за минуту, известно как доставка кислорода и равняется сердечному выбросу х содержание кислорода в артериальной крови (см. выше). Например, 5000 мл крови/мин х 200 мл О2/1000 мл крови = 1000 мл О2/мин. Доставка кислорода (млО2/мин) = сердечный выброс (л/мин) х концентрация Нb (г/л) х 1,30 (млО2/г Hb) х % сатурации.
Рис. 3. Кислородный каскад.
Эффекты гиповентиляции показаны тонким пунктиром, шунта — ширным
Человеком в покое каждую минуту потребляет около 250 мл кислорода (потребление кислорода), то есть используется всего 25% кислорода артериальной крови. Гемоглобин в смешанной венозной крови насыщен примерно на 75% (95% — 25%).
Клеткам доставляется больше кислорода, чем они могут использовать. При высоком потреблении кислорода (например, при физической нагрузке) повышенная потребность компенсируется повышением сердечного выброса (см. формулу выше). Однако низкий сердечный выброс, низкое содержание гемоглобина (анемия) или низкая сатурация гемоглобина приведет к неадекватной доставке кислорода, если не произойдет компенсаторных изменений в одном из перечисленных звеньев. С другой стороны, если доставка кислорода падает на уровень потребления кислорода, ткани начинают экстрагировать большее его количество (сатурация смешанной венозной крови падает ниже 70%)(рис. 4 а-b). Снижение доставки ниже точки «с» на рисунке 4 не может быть компенсировано повышением экстракции кислорода и ведет к анаэробному метаболизму и лактат ацидозу.
Несмотря на огромную важность кислорода, его запасы в организме человека невелики и недостаточны для поддержания жизни более, чем несколько минут. При отсутствии дыхания запасы кислорода ограничены его наличием в легких и крови. Количество кислорода в рови зависит от объема циркулирующей крови и концентрации Hb. Количество в легких – от величины функциональной остаточной емкости (ФОЕ) и альвеолярной концентрации кислорода. ФОЕ это объем воздуха (около 3 литров у взрослых), который остается в легких в конце выдоха, когда эластические силы легких находятся в равновесии с диафрагмой и расслабленной грудной клеткой. При дыхании общий запас кислорода (кислород крови и легких) невелик и так как в основном он связан с гемоглобином (см. рис. 5), только небольшая его часть высвобождается при отсутствии недопустимого падения РО2 (когда насыщение — сатурация гемоглобина 50% и РО2 снижено до 26 мм Hg). Дыхание 100% кислородом приводит к значительному увеличению запасов кислорода в организме, так как ФОЕ заполнена кислородом. Основной компонент запаса теперь находится в легких и 80% его может быть использовано без опасности критического снижения сатурации гемоглобина (РАО2 остается около 100 ммHg). Это объясняет эффективность преоксигенации (см. ниже).
Транспорт кислорода – эффекты анестезии
Гиповентиляция может возникать во время анестезии вследствие обструкции дыхательных путей, действия ингаляционных анестетиков, опиоидов и гипнотиков. С другой стороны, при анестезии кетамином и эфиром (менее 1 МАК) наблюдается менее выраженная депрессия дыхания, чем при использовании других анестетиков. Альвеолярное РО2 сбалансировано поступлением кислорода при дыхании и его потреблением в метаболических процессах организма. Гиповентиляция и сниженная вдыхаемая концентрация кислорода приведет к падению альвеолярного РАО2. Повышенное потребление кислорода при растущих метаболических потребностях, например, при послеоперационном ознобе или злокачественной гипертермии также приводит к снижению РaО2.
При падении РаО2 ниже 60 мм Hg хеморецепторы каротидного синуса и дуги аорты вызывают гипервентиляцию и повышение сердечного выброса через стимуляцию симпатической нервной системы. Эту нормальную протективную реакцию на гипоксию подавляют анестетики в интра- и послеоперационном периоде.
После индукции в наркоз наблюдается быстрое снижение ФОЕ, приводящее с закрытию мелких бронхиол, особенно в зависимых частях легких, которые спадаются и находятся в закрытом состоянии втечение всего респираторного цикла. Некоторые альвеолы при этом не вентилируются и возникает шунт.
Нарастают V/Q нарушения, что приводит к снижению сатурации крови в легочных венах ниже, чем в капиллярах вентилируемых альвеол. Ситуация усугубляется во время индукции в наркоз, когда примесь неоксигенированной крови может составлять не 1%, а 10%. За исключением пациентов с сохраненным спонтанным дыханием на фоне анестезии кетамином подобная картина возникает вне зависимости от использованного анестетика и от того, вводились ли пациенту миорелаксанты. Это следует рассматривать как неизбежный побочный эффект анестезии. Ингаляционные анестетики подавляют легочную гипоксическую вазоконстрикцию и кровоток по невентилируемым альвеолам не снижается. Многие анестетики снижают сердечный выброс и соответственно доставку кислорода. При анестезии на 15% падает уровень метаболизма и, следовательно, потребность в кислороде. Искусственная вентиляция снижает потребности в кислороде на 6% вследствие отсутствия работы дыхательной мускулатуры. Анестетики не влияют на кислородотранспортные свойства гемоглобина.
Рис. 4. На рисунке горизонтальной линией показан лимит, до которого снижение доставки О2 не вызовет снижение его потребления (оксигенация, не зависимо от доставки) (А-В). Ниже указанного критического значения доставки О2 потребление становится зависимым от доставки и возникают признаки гипоксии. Положение точки С зависит от значения максимально возможной экстракции кислорода.
Читайте также: Ткань кашемир 672 мебельная
Из протекающей артериальной крови ткани усваивают 6 кислорода
В сложных механизмах транспорта газов кровью и газообмена в тканях важная роль отводится эритроцитам, ответственным за доставку О2 к различным органам и удаление образующегося в процессе метаболизма СО2.
Эритроцит – безъядерная клетка, лишенная митохондрий, основным источником энергии для эритроцита служит глюкоза, метаболизируемая в гексозомонофосфатном шунте или цикле Эмбдена-Мейергофа. Транспорт О2 обеспечивается в значительной мере гемоглобином, состоящим из белка глобина и гема. Последний представляет собой комплексное соединение железа и порфирина. Глобин представляет собой тетрамер полипептидной цепи. Hb A (HbA) – основной гемоглобин взрослых содержит 2 – альфа и 2 – бета – цепи, Hb A2 – содержит две альфа и две дельта цепи.
Гем состоит из иона железа, встроенного в порфириновое кольцо. Ион железа гема обратимо связывает одну молекулу О2. С одной молекулой Hb максимально связываются 4 молекулы О2 с образованием оксигемоглобина.
Гем может подвергаться не только оксигенации, но и истинному окислению, когда железо становится из двухвалентного трехвалентным. Окисленный гем носит название гематина, а молекула гемоглобина становится метгемоглобином. В крови человека метгемоглобин находится в незначительных количествах, его уровень резко возрастает при отравлениях. Метгемоглобин не способен отдавать кислород тканям.
В норме метгемоглобин составляет менее 3% общего Hb крови. Основная форма транспорта О2 – в виде оксигемоглобина. Кислород транспортируется артериальной кровью не только в связи с гемоглобином, но и в растворенном виде. Принимая во внимание тот факт, что 1 г Hb может связать 1,34 мл О2, кислородная емкость крови в среднем у взрослого человека составляет около 200 мл/л крови. Одним из показателей кислородного транспорта является насыщение артериальной крови О2(Sa O2), равного отношению О2, связанного с Hb, к кислородной емкости крови:
SaO2=O2, связанного с Hb/O2 емкость крови* 100%.
В соответствии с кривой диссоциации оксигемоглобина насыщение артериальной крови кислородом в среднем составляет 97%, в венозной крови – 75%.
PaO2 в артериальной крови около 100 мм. рт. ст., а в венозной – около 40 мм. рт. ст.
Количество растворенного кислорода в крови пропорционально парциальному давлению О2 и коэффициэнту его растворимости.
Последний для О2 составляет 0,0031/100 мл крови/ 1 мм. рт. ст.. Таким образом, 100 мл крови при PaO2, равном 100 мм. рт. ст., содержит менее 0,31 мл O2.
Диссоциация оксигемоглобина в тканях обусловлена главным образом химическими свойствами гемоглобина, а также рядом других факторов – температурой тела, рН среды, р СО2.
При понижении температуры тела наклон кривой диссоциации оксигемоглобина возрастает, а при ее повышении – снижается, и соответственно снижается сродство Hb к О2.
При снижении рН, т.е. при закислении среды, сродство гемоглобина к О2 уменьшается. Увеличение напряжения в крови СО2 также сопровождается снижением сродства Hb к О2 и уплощением кривой диссоциации оксигемоглобина.
Известно, что степень диссоциации оксигемоглобина определяется содержанием в эритроцитах некоторых фосфорорганических соединений, главным из которых является 2,3 – ДФГ (2,3 дифосфоглицерат), а также содержанием в эритроцитах катионов. В случаях развития алкалозов, поглощение О2 в легких увеличивается, но в то же время затрудняется отдача кислорода тканями. При ацидозах наблюдается обратная картина.
4.2.Утилизация кислорода тканями
Тканевое или клеточное дыхание включает три стадии. На первой стадии пируват, аминокислоты и жирные кислоты окисляются до двухуглеродных фрагментов ацетильных групп, входящих в состав ацетилкофермента А. Последние на втором этапе окисления включаются в цикл лимонной кислоты, где происходит образование высокоэнергетических атомов водорода и высвобождение СО2 – конечного продукта окисления органических субстратов. На третьей стадии клеточного дыхания атомы водорода делятся на протоны (Н+) и «высокоэнергетические» электроны, передающиеся по дыхательной цепи на молекулярный О2 и восстанавливающие его до НО2. Перенос электронов сопряжен с запасом энергии в форме АТФ, т.е. с окислительным фосфорилированием (рис.6).
Касаясь патогенеза метаболических сдвигов, свойственных гипоксическим состояниям, следует отметить, что в организме человека более 90% всего потребляемого кислорода восстанавливается с участием цитохромоксидазы митохондрий, и лишь около 10% кислорода метаболизируется в тканях с участием оксигеназ: диоксигеназы и монооксигеназы.
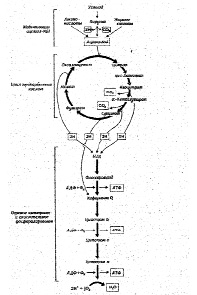
Рис.6. Схема тканевого дыхания. Конечные продукты каждой стадии даны в рамке (Ленинджер А., 1999)
Наиболее многочисленны и сложны монооксигеназные реакции, протекающие в эндоплазматическом ретикулуме клеток при участии цитохрома Р-450 и обеспечивающие гидроксилирование субстрата (стероидных гормонов, лекарственных препаратов и различных др. соединений) и, как правило, его инактивацию.
Диоксигеназы катализируют реакции, в которых в молекулу органического субстрата включаются оба атома молекулы кислорода (например, реакция окисления катехола молекулярным кислородом с раскрытием кольца).
Читайте также: Парка из мембранной ткани женская
В реакциях, связанных с переносом электронов, т.е. в реакциях окисления-восстановления, где, как указывалось выше, используется более 90% потребляемого кислорода, атомы водорода, отщепленные дегидрогеназами от субстратов в цикле лимонной кислоты, передают свои электроны в цепь переноса электронов и превращаются также в Н +. Как известно, помимо 4 пар атомов водорода, поставляемых каждым оборотом цикла лимонной кислоты, образуются и другие атомы водорода, отщепленные дегидрогеназами от пирувата, жирных кислот и аминокислот в процессе их расщепления до Ацетил-СоА и других продуктов.
Таким образом, все атомы водорода, отщепляемые дегидрогеназами от субстратов, передают свои электроны в дыхательную цепь к конечному акцептору электронов – кислороду.
Касаясь последовательности транспорта электронов в окислительно-восстановительных реакциях, протекающих на внутренней мембране митохондрий, прежде всего, следует отметить, что от всех НАД – зависимых реакций дегидрирования восстановленные эквиваленты переходят к митохондриальной НАДН – дегидрогеназе, затем через ряд железосерных ферментов передаются на убихинон М цитохрому b. Далее электроны переходят последовательно на цитохромы С1 и С, затем на цитохромы аа 3 (цитохромоксидазу – медьсодержащий фермент). В свою очередь цитохромоксидаза передает электроны на кислород. Для того, чтобы полностью восстановить кислород с образованием 2-х молекул воды требуется 4 электрона и четыре Н+ .
Скорость утилизации О2 в различных тканях различна. В среднем взрослый человек потребляет 250 мл О2 в 1 мин. Максимальное извлечение О2 из притекающей артериальной крови свойственно миокарду.
Кислород используется в клетках, в основном в метаболизме белков, жиров, углеводов, ксенобиотиков, в окислительно-восстановительных реакциях в различных субклеточных фракциях: в митохондриях, в эндоплазматическом ретикулуме, в реакциях липопероксидации, а также в межклеточном матриксе и в биологических жидкостях.
Коэффициент утилизации О2 в тканях равен отношению потребления О2 к интенсивности его доставки, широко варьирует в различных органах и тканях.
В условиях нормы минимальную потребность в О2 проявляют почки и селезенка, а максимальную потребность – кора головного мозга, миокард и скелетные мышцы, где коэффициент утилизации О2 колеблется от 0,4 до 0,6, а в миокарде до 0,7. При крайне интенсивной физической работе коэффициент утилизации О2 мышцами и миокардом может возрастать до 0,9.
Обмен дыхательных газов в тканях происходит в процессе свободной и облегченной диффузии. При этом О2 переносится по градиенту напряжения газа из эритроцитов и плазмы крови в окружающие ткани.
Одновременно происходит диффузия СО2 из тканей в кровь. На выход О2 из крови в ткани влияет диссоциация оксигемоглобина в эритроцитах, что обеспечивает так называемую облегченную диффузию О2. Интенсивность диффузионного потока О2 и СО2 определяется градиентом их напряжения между кровью и тканями, а также площадью газообмена, плотностью капилляров, распределением кровотока в микроциркуляторном русле. Интенсивность окислительных процессов в тканях определяется величиной критического напряжения О2 в митохондриях, которое в условиях нормы должно превосходить 0,1-1 мм рт. ст.
Соответствие доставки О2 к органам и тканям, возросшим потребностям в оксигенации обеспечивается на клеточном, органном уровнях за счет образования метаболитов изнашивания, а также при участии нервных, гормональных и гуморальных влияний.
Основная масса углекислого газа (СО2) образуется в организме как конечный продукт различных метаболических реакций и транспортируется к легким с кровью. Вдыхаемый воздух содержит лишь незначительное количество СО2.
Транспорт СО2 кровью осуществляется в 3-х состояниях: в виде аниона бикарбоната, в растворенной форме и в виде карбаминовых соединений.
СО2 хорошо растворяется в плазме крови и в артериальной крови, около 5% от общей двуокиси углерода содержится в крови в растворенной форме.
Анион бикарбоната составляет около 90% от общего содержания СО2 в артериальной крови: СО2+Н2О — Н++НСО-3.
Эта реакция медленно протекает в плазме крови, но чрезвычайно интенсивно происходит в эритроцитах при участии фермента карбоангидразы. Мембрана эритроцита относительно непроницаема для Н+, как и вообще для катионов, но в тоже время, проницаема для ионов НСО-3, выход которых из эритроцитов в плазму обеспечивается притоком Cl- из плазмы в эритроциты. Часть Н+ забуферивается гемоглобином с образованием восстановленного гемоглобина.
Третьей формой транспорта СО2 кровью являются карбаминовые соединения, образованные взаимодействием СО2 с концевыми группами белков крови преимущественно с гемоглобином:
Hb NH2 + CO2 — Hb NH COOH > Hb NH COO + Н+
Карбаминовые соединения составляют около 5% от общего количества СО2, транспортируемого кровью.
В оксигенированной артериальной крови напряжение СО2 составляет 40 мм. рт. ст., а в венозной крови Рv СО2 равно 46 мм. рт. ст.
4.4.Связывание гемоглобина с окисью углерода
Угарный газ (СО) – окись углерода обладает значительно большим сродством к гемоглобину, чем О2, с последующим образованием карбоксигемоглобина. СО входит в состав бытового газа, а также выделяется при работе двигателей внутреннего сгорания. При концентрации СО во вдыхаемом воздухе всего в количестве 7*10- 4 около 50% гемоглобина превращается в карбоксигемоглобин. В норме в крови содержится около 1% HbCO, у курильщиков – 3% . В крови водителей такси концентрация карбоксигемоглобина достигает 20%. Карбоксигемоглобин диссоциирует с отдачей О2 в 200 раз медленней оксигемоглобина и в то же время препятствует его диссоциации в тканях.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом